ЭКОЛОГО-ГИДРОГЕОЛОГИЧЕСКОЕ ПРЕДПРИЯТИЕ
"ЭКОМОНИТОРИНГ"
СРЕДНЕУРАЛЬСКАЯ ГИДРОГЕОЛОГИЧЕСКАЯ ЭКСПЕДИЦИЯ
|
620014
г.Екатеринбург ул.Вайнера, 55 (Уралнедра), каб. 513 |
тел. 257-20-06, 219-39-08 факс 257-20-06 |
|
|
|
Глава шестая. Происхождение и развитие планеты Земля Возникновение
Земли Земная
кора, или литосфера. Хотя есть основания считать, что возраст Земли составляет
около 4,5 миллиардов лет, в вопросе о том, как образовалась наша планета и
вся Солнечная система, частью которой является Земля, еще нет ясности. Все,
чем мы располагаем, - это теория, которая постоянно меняется по мере
сопоставления с вновь получаемыми нами сведениями о Солнечной системе. Основные
положения этой теории сводятся к тому, что вся Солнечная система - Земля,
другие планеты и Солнце - образовалась одновременно и как единое целое. Такой
процесс образования системы не представляет собой что-то уникальное или хотя
бы необычное, а является вполне заурядным. Подобные процессы происходят во
Вселенной и в настоящее время. Исходным материалом для образования Солнечной
системы явилась туманность - огромное вращающееся дискообразное
облако, состоящее из холодного газа и "пыли". Из этой холодной и
распыленной массы по мере ее уплотнения и выделялись группы отдельных
сгущений, из которых в конце концов образовались Солнце и планеты. Одно из
этих сгущений, которое позднее стало Землей, имеющей ныне диаметр 13 400
километров, первоначально имело диаметр в несколько миллионов километров,
настолько далеко друг от друга находились составляющие его частицы. Но эти
частицы постепенно приближались друг к другу под действием сил тяготения и
теряли энергию в результате бесчисленных столкновений. Более крупные частицы
"падали", подобно градинам, в направлении центра сгущения, и при
этом гравитационном сжатии выделялось такое большое количество тепла, что в
итоге происходило резкое уплотнение массы с образованием твердого тела. Твердое
тело Земли первоначально было очень горячим (То есть, возможно, было
разогрето по мере образования Земли теплом радиоактивного распада и
гравитацией. - Прим. ред.). Предполагается, что частицы железа, входившие в
состав этой юной, горячей, уплотняющейся Земли, плавились и образовывали
тяжелые капли, которые опускались к центру Земли. Там происходило накопление
железа и других минералов, в результате чего было создано ныне существующее
ядро Земли. Более
легкие частицы, "падавшие" не глубоко, образовали толстый слой,
окруживший более тяжелое ядро. Этот слой был в большей степени каменным, чем
металлическим, и состоял из силикатных минералов. Так образовалось то, что мы
называем мантией; о ней уже говорилось в первой главе. Толстый
слой силикатов, плохо проводящий тепло, препятствовал, как и в данное время,
оттоку тепла от нагретой внутренней части Земли. Но, хотя температура мантии
стала высокой, плавления в ее глубоких частях не происходило, так как вес
вышележащих толщ создавал слишком высокое давление. Но в верхней (или
внешней) части мантии, где слой перекрывающих пород был значительно тоньше и
давление меньше, породы частично плавились. Правда, плавились только минералы
с наинизшей точкой плавления, другими словами,
плавление было избирательным. Оно и создавало магму. Поскольку магма
образовалась лишь из некоторых минералов окружающей породы, то по составу она
отличалась от этой исходной породы. Состав ее приближался к базальтовому
слою, и вес был немного меньше, чем у окружающей нерасплавленной породы. Более
легкие массы вновь образованной базальтовой магмы имели тенденцию к поднятию.
Очагов таких масс было очень много, и поднятие их происходило медленно.
Постепенно они затвердевали на поверхности Земли, где температуры были более
низкими. По мере затвердевания постепенно образовывалась кора, состоящая из
базальтов и покрывающая всю Землю. Таким образом, базальтовая кора является
продуктом избирательного плавления, и ее состав несколько отличается от
состава мантии. Это избирательное плавление некоторых компонентов первичных
горных пород, вероятно, представляло собой первый процесс длинной цепи
процессов постепенной дифференциации первичной магмы и формирования горных
пород, слагающих земную кору. Вторым
процессом было химическое выветривание, третьим - образование гранитных
пород, которые легче базальта. Очевидно, все эти три процесса идут до сих
пор. Атмосфера
и гидросфера. До сих пор мы излагали теорию образования внешней части
литосферы. А как образовались атмосфера и гидросфера? Хотя на первый взгляд
это кажется странным, обе эти подвижные оболочки образовались в основном тем
же способом, что и твердая кора: путем поднятия магмы к поверхности (или на
поверхность) Земли. Это оказалось возможным потому, что всякая магма содержит
газы. Выше мы сказали, что магма образовалась местами в результате плавления
глубинной породы под действием тепла, поступавшего из нагретых внутренних
частей. В процессе плавления некоторые минералы легко разлагались на
составные химические элементы. Этот процесс отличался от сортировки
кристаллов минералов по весу. Распад вещества происходил под действием
температуры, давления и химически активной среды в магме. Среди других
веществ, высвобождавшихся таким образом, были водород (Н2),
азот (N2), окись углерода (СО), двуокись
углерода (СО2) и вода (Н2О). Поднимающаяся магма выносила эти
вещества на поверхность, и там они из нее выделялись. Таким образом началось
создание атмосферы Земли, атмосферы, которая на этих ранних этапах была
целиком вулканической. Она состояла из метана, водяного пара, аммиака,
водорода, азота, и, возможно, некоторого количества окиси и двуокиси
углерода. Те же элементы и вещества содержатся в газах, выделяемых
современными вулканами. Наиболее существенным отличием древней атмосферы от
современной было то, что весь кислород в ней был связан в различных
химических соединениях. Свободного кислорода не было. Это очень важно, так
как бедная кислородом атмосфера оказывает большое влияние на образование
живого вещества, как мы увидим в седьмой главе. Что
касается гидросферы, то не трудно понять, что она образовалась из атмосферы,
содержавшей большое количество водяного пара, поступавшего из вулканов.
Водяной пар конденсировался и выпадал в виде дождя. При стоке дождевой воды формировались
потоки, заканчивавшиеся в озерах, занимавших понижения земной поверхности.
Озера росли и сливались, образуя океан. В ходе этого процесса начался
круговорот воды. При
соприкосновении атмосферы, содержавшей углекислый газ и воду, с молодыми базальтовыми
породами, выходящими на поверхность, должно было начаться их выветривание.
Вновь образующиеся потоки несли продукты выветривания, частицы пород и
растворенные вещества в новые озера. Веществом, растворенным в воде растущих
и затопляющих поверхность суши озер, была соль (на самом деле смесь многих
солей), присутствовавшая в воде зарождавшегося океана. Так было положено
начало засолению морской воды, которое мы наблюдаем сейчас. Хотя
такая последовательность событий представляется правдоподобной, потому что
она хорошо согласуется с нашими знаниями о Земле, все же это только теория, и
далеко не доказанная. Кроме того, очень мало известно, сколько времени все
это продолжалось - сколько времени потребовалось для того, чтобы атмосфера и
океан приобрели свой современный состав и размеры. Есть основания полагать,
что в течение по крайней мере последних полмиллиарда или миллиарда лет
существенных изменений в составе этих подвижных оболочек не произошло. Таким
образом, представляется вероятным, что составные части земной коры были
созданы в ходе вулканической деятельности, проявлявшейся одновременно с
выветриванием пород, опусканием и повторным расплавлением пород на большой
глубине - иначе говоря, протекавшей одновременно с круговоротом вещества.
Вероятно также, что составные части атмосферы и гидросферы
"испарились" понемногу из горячих глубин Земли. Они поднялись вверх
в виде газов, растворенных в поднимающейся магме, и выделились при
извержениях вулканов. Такой перенос вверх происходил начиная с самых ранних
этапов истории Земли, и до сих пор на поверхность Земли поступает
"новая" вода. К настоящему времени большая часть воды, выделяемой
вулканами, - это "старая" вода, которая многократно находилась в
круговороте и была нагрета вулканическим теплом, будучи в толще Земли. Но
если только полпроцента воды, выделяемой вулканами, представляет собой
"новую" воду, то, согласно расчетам, при такой скорости поступления
воды на поверхность вся гидросфера, включая океан и все остальное, могла бы
быть создана за четыре миллиарда лет. Эта
теория происхождения земной коры, водной оболочки и газовой оболочки Земли
очень хорошо согласуется с принципом актуализма, которому подчинены все
геологические процессы. Отсюда следует вывод о том, что даже в наиболее
отдаленные времена действовали те же процессы, что и сейчас, и характер их
проявления был во многом сходен с современным. Луна.
Характер и история Луны имеют связь с нашей теорией происхождения Земли.
Данные о Луне, полученные благодаря программе "Аполлон", начатой в
1969 г., до сих пор не выявили ничего не сопоставимого с представлениями об
истории Земли, созданной по геологическим данным. По всей вероятности, Луна,
очень малый член Солнечной системы, была образована таким же путем, как и
Земля, и независимо от Земли. Представляется, что по крайней мере ее внешняя
часть подвергалась когда-то расплавлению. Найденные там породы большей частью
темноокрашенные магматические породы, частично базальты, частично породы,
подобные габбро. Это свидетельствует о поднятии легких компонентов к
поверхности в процессе застывания, в то время как более тяжелые компоненты
опускались вниз. Мы думаем, что по крайней мере частично это происходило и
при образовании земной коры. Но
на этом сходство с Землей кончается. Луна покрыта слоем (толщиной 1,5-3
метра) вещества, обычно называемого "лунной пылью". Мы зовем его
лунным реголитом. Он состоит из продуктов механического разрушения
нижележащих коренных пород. Некоторые из них представляют собой большие или
малые обломки, но большая часть пылевидная. Такое механическое разрушение
вызвано не физическим выветриванием, как на Земле, - это результат
бомбардировки малыми и большими метеоритами лунной поверхности. Некоторые из
них достигали огромной величины. Удары метеоритов вызвали также и образование
лунных кратеров. Конечно, метеориты попадают и в атмосферу Земли, самое
меньшее 20 миллионов ежедневно. Но большей частью они очень малы и
"сгорают" - распадаются - в результате трения при прохождении через
атмосферу. Время от времени крупные метеориты попадают и на поверхность
Земли, и когда это случается, в месте падения образуется кратер. Лунная
пыль - реголит Луны - не обнаруживает никаких следов химического
выветривания, обычных для рыхлого чехла земных пород. Причиной этого является
отсутствие на Луне атмосферы и гидросферы. Отсутствие это нетрудно объяснить.
Земля имеет достаточно большую массу, чтобы сила ее тяготения удерживала
газы, составляющие ее атмосферу. Но масса Луны составляет только 0,012 массы
Земли и недостаточна для удержания каких-либо газов. Поэтому на поверхности
Луны не происходит химическое выветривание. Континенты
и плиты коры В
первой главе отмечалось, что континентальная кора Земли гораздо толще, чем
океаническая, и отличается от последней по составу. В пятой главе было
рассказано, как отдельные континенты медленно увеличивают свои размеры. .Наша
карта современной поверхности Земли отличается от карт поверхности Земли,
существовавшей, скажем, сто миллионов лет назад. Такие большие изменения
трудно понять, если не представить себе сначала общую картину строения Земли. Континенты
не остаются на месте, а движутся. Достойно изумления, что впервые
предположение о таком движении было выдвинуто около 350 лет назад, и с тех
пор выдвигалось еще несколько раз, однако эта идея получила признание ученых
только после 1960 г. Большинство людей считало, что жесткость коры исключает
движение континентов. Теперь все мы знаем, что это не так. Чтобы понять, что
происходит с континентами, давайте рассмотрим некоторые доказательства их
движения. Доказательства
движения континентов. Доказательства того, что по крайней мере некоторые
континенты, разобщенные сейчас, раньше соединялись, или того, что они
изменили положение, включают следующее: 1.
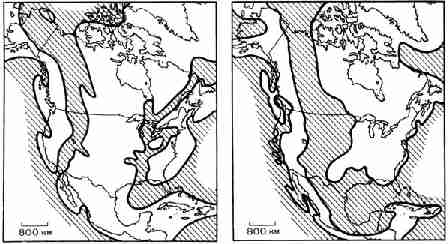
Контуры некоторых континентов, обращенных друг к другу и разделенных океаном,
позволяют предположить, что в прошлом они соединялись, как две части
головоломки. Наиболее очевидный пример этого - Южная Америка и Африка. Так
как истинной окраиной континента является не современная береговая линия, а
скорее внешний край континентального шельфа, проверять соответствие
континентов лучше не по обычной карте, а по карте, показывающей шельфы (рис.
16). 2.
На некоторых парах континентов обнаруживается соответствие другого рода -
особые типы пород, древние горные пояса, одинаковые ископаемые животные и
растения. Все эти совпадения позволяют предполагать, что такие континенты
когда-то соединялись. 3.
Ископаемые животные и растения, которые должны были жить в теплом климате, встречаются
на некоторых континентах, на таких широтах, где современный климат гораздо
холоднее. 4.
Убедительные доказательства широкого распространения древних ледников
встречаются в породах, возраст которых около 250-300 миллионов лет, в
восточной части Южной Америки, в Южной Африке, Индии, Австралии, на таких
широтах, где сейчас теплый климат препятствует образованию ледников. Подобные
следы древнего оледенения найдены в Антарктиде, где ледники существуют и
ныне. Представляется, что все эти континенты были некогда частями одного
континента, который распался на куски. Они двигались порознь. Некоторые из
них переместились в более теплые климатические пояса, а Антарктида заняла
свое исключительное положение на Южном полюсе. 5.
Во многих пластах содержатся свидетельства того, что континенты переместились
по отношению к магнитному Северному полюсу. Это положение требует
разъяснения. Магнитные
данные. Если мы хотим знать точное направление, мы пользуемся компасом.
Стрелка, будучи намагниченной, располагается вдоль линий магнитного поля
Земли; она указывает на магнитный Северный полюс. Подобно этому, некоторые
горные породы являются естественными компасами. Переносящиеся водным потоком
кристаллы магнитных железистых минералов, соответствующие по размеру частицам
песка или алеврита, ведут себя подобным же образом. Когда они осаждаются в
русле, они ориентируются вдоль линий север - юг, и во многих случаях эта
ориентировка сохраняется, даже когда осадок в конце концов становится
осадочной породой. Подобное
воздействие претерпевают магнитные железистые минералы в магматических
породах, например, в базальтах. Когда магма или лава остывает, она достигает
такой температуры, ниже которой эти минералы могут приобрести
намагниченность, параллельную магнитному полю Земли. Эта магнитная
ориентировка (не имеющая ничего общего с ориентировкой самого кристалла в
породе) сохраняет ориентировку магнитного поля, существовавшего тогда, когда
остывали породы. Такие железистые минералы представляют собой как бы стрелки
компасов, указывающие положение магнитного полюса. Ученые
обнаружили указатели прежнего направления магнитного поля в слоях различного
возраста и не поленились измерить их в сотнях точек и в целой серии различных
стратиграфических горизонтов (соответствующих серии различных моментов
времени в прошлом). Результирующие направления отличаются от тех, которые
компас показывает в этих же точках в настоящее время. Подобные направления
свидетельствуют, что по крайней мере за последние 200 миллионов лет или около
того континенты испытали значительные перемещения не только по отношению к
полюсу, но и по отношению друг к другу. Путь, пройденный тем или другим
континентом, может быть прослежен, по крайней мере в общих чертах, если не
вполне точно. В
ходе исследований ориентировки древнего магнитного поля выявился
поразительный факт. Было обнаружено, что во многих слоях Земли, накопившихся
за последние несколько миллионов лет (промежуток времени, охваченный
исследованиями), намагниченность частиц обратна
современной. Магнитные
частицы в породах указывают в сторону не Северного, а Южного полюса.
Радиометрические датировки тех же пластов показывают, что обратная
ориентировка фиксируется на всем земном шаре в одни и те же промежутки
времени. Очевидно, в это время само магнитное поле Земли, контролирующее эту
ориентировку, имело обратное направление, и два его полюса менялись местами.
Такие "обращения" магнитного поля происходили неоднократно. Что
заставляло магнитное поле изменять направление? Мы пока не знаем, но
предполагается, что изменения могли быть вызваны движениями в жидкой части
железосодержащего ядра Земли. Во всяком случае, они проявлялись на всей Земле
сразу. Оказалось
возможным построить календарь изменений магнитного поля для всего земного
шара. Этот календарь (рис. 20) показывает промежутки времени, в течение
которых магнитное поле было нормальным (таким, как в нынешнее время), и
промежутки, когда оно было обращенным. Обращения (инверсии) магнитного поля
Земли, открытие которых сделало возможным создание указанного календаря, представляют
собой явление, совершенно отличное от колебаний ориентировки магнитного поля,
измерения которых легли в основу вывода о движении континентов.
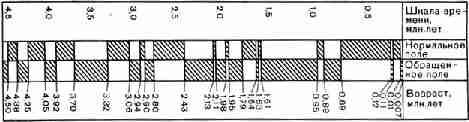
Рис. 20. Хронологическая шкала обращений магнитного
поля Земли, построенная по данным радиометрических определений возраста
пород, для которых измерялось направление намагниченности Сравнивая
магнитный календарь с геохронологической шкалой (табл. 1), можно увидеть, что
отрезок времени длиной в 4,5 миллиона лет, который охватывает этот календарь,
включает весь голоцен, весь плейстоцен и часть плиоцена. Вооруженные знанием
того, что магнитные обращения случаются и запечатлеваются в ориентировке
магнитного ноля в затвердевших базальтах на континентах, мы можем теперь
воспользоваться этим при рассмотрении магнитной ориентировки базальтов на дне
океана. Срединные
океанические рифты. Самым замечательным в серии открытий, сделанных после
1950 г. в изучении океана, было признание того, что дно океана не
представляет собой плоскую монотонную равнину; формы его так же разнообразны,
как и формы поверхности континентов. Среди многих форм есть и глубокий рифт -
долина, продолжающаяся почти непрерывно на протяжении более 6000 километров,
местами глубиной более 1000 метров, которая делит на две части Атлантический
океан (рис. 22) и пересекает Индийский и Тихий океаны.
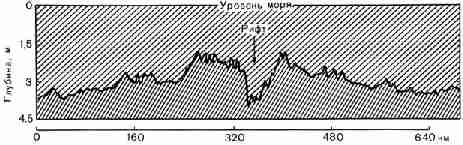
Рис. 21. Широтный профиль дна средней части
Атлантического океана. Виден рифт (в данном месте глубиной около 1800
метров), по которому происходит поднятие базальтовых лав Рифт
занимает центральную, наиболее приподнятую часть срединного океанического
хребта. Это странное соотношение отчетливо видно на рисунке 21. Рифт гораздо
глубже глубокой долины, изображенной на этом рисунке, он продолжается под
поверхностью на 50-100 километров в глубь Земли, и по всей его длине на
поверхность дна океана поступает огромное количество тепла из глубин.
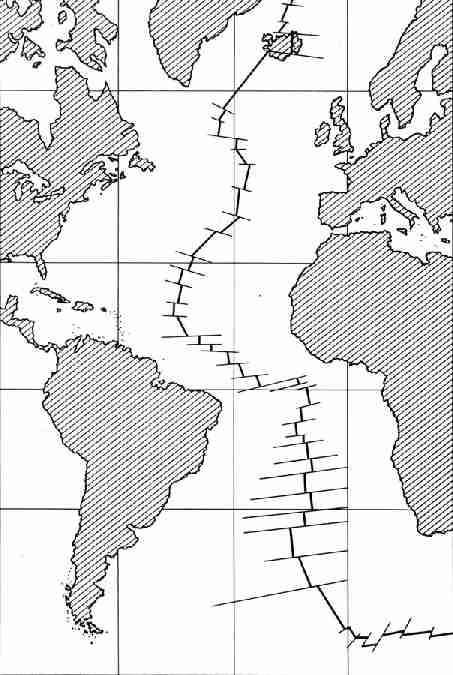
Рис. 22. Рифт, разделяющий на две части дно
Атлантического океана. Он разбит на отдельные фрагменты, смещенные влево или
вправо по поперечным линиям, представляющим собой разломы, которые
продолжаются в глубь земной коры. Продолжение рифта за пределами
Атлантического океана на рисунке 25 Под
тонким покровом рыхлых осадков, выстилающих большую часть дна глубоководных
областей океана, по обеим сторонам рифта ложе океана сложено базальтами.
Когда исследовательские корабли, оборудованные магнитометрами, начали
производить измерения магнитной полярности базальтов, слагающих дно океана на
глубине несколько километров, курсируя в западном и восточном направлениях
поперек зоны рифта на сотни километров, магнитометрическая съемка дала
поразительные результаты. Оказалось, что на дне выявляются чередующиеся
полосы, или зоны нормальной (северной) и обращенной (южной) намагниченности.
Эти полосы параллельны рифту, причем на различных сторонах рифта они
представляют зеркальное отображение (рис. 23). Последовательность узких и
широких полос совпадает с верхней частью календаря изменений магнитного поля
Земли, изображенного на рисунке 20. Основываясь на этом совпадении, возраст
каждой полосы можно считать таким же, как возраст соответствующего
подразделения указанного календаря. Самые молодые полосы базальта залегают
непосредственно вблизи рифта, к ним примыкают более древние. С удалением от
рифта вправо и влево мы встречаем все более и более древние полосы.
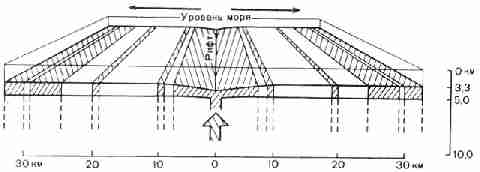
Рис. 23. Схема, показывающая полосчатое строение
базальтов, образующихся в зоне срединно-океанического рифта при затвердевании
поднимающейся магмы (показанной широкой стрелкой). Базальт постоянно
перемещается вправо и влево от зоны рифта (показано тонкими стрелками) по
мере наращивания Дна океана. Каждая полоска сохраняет направление магнитной
полярности, приобретенное ею при затвердевании. Штриховкой показаны полосы с
нормальной намагниченностью, светлые полосы имеют обращенную намагниченность Эта
поразительная закономерность может обозначать лишь одно обстоятельство.
Очевидно, происходит непрерывное образование новой земной коры в результате
затвердевания лавы, поднимающейся из глубин в зоне рифта. Кора должна
перемещаться из области рифта в противоположных направлениях, часть к западу,
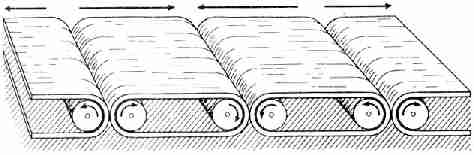
а часть к востоку, как два конвейера, движущиеся наружу от общей оси (рис.
24). Вновь образующаяся лава сохраняет нормальную или обращенную полярность,
которая запечатлевается в ней в результате воздействия магнитного поля Земли
в момент остывания. Каждый раз, когда поле Земли меняет ориентировку на
обратную, начинается новая зона. Происходит это потому, что зоны выделяются
не по составу лавы (которая, постоянно поднимаясь и застывая, всегда имеет
одинаковый состав), но исключительно по полярности зерен магнитных минералов,
содержащихся в ней. Другими словами, без магнитометрической съемки мы не
могли бы выделить этих полос.
Рис. 24. Серия из четырех конвейеров, движущихся во
взаимно противоположных направлениях. Сравните рис. 23 Какова
же скорость этого необычайного процесса? Современная скорость рассчитывается
по шкале магнитных обращений (рис. 20), и оказалось, что кора в западной
части Атлантического океана движется сейчас на северо-запад со скоростью 1-2
сантиметра в год. Если
мы примем скорость постоянной и равной 1,5 см/год, то одно только перемещение
этой части должно было расширить Атлантический океан за последние 100
миллионов лет на 1500 километров. Движение восточной окраины рифта на восток
должно было еще больше увеличить ширину Атлантики. Действительно, имеются
доказательства того, что 150-200 миллионов лет назад (то есть в,
позднетриасовое, или раннеюрское, время) Атлантический океан не существовал,
а восточный берег Северной Америки, имел жесткое соединение с северо-западной
Африкой. Однако
кора в области Атлантики движется сравнительно медленно. Расчеты с помощью
шкалы обращений магнитного поля Земли показывают, что в Индийском океане
скорость движения достигает 3 см/год, а в некоторых частях Тихого - 6 см/год
и более. Ширина черных полос на рисунке 25 показывает площади
новообразованной коры вдоль срединно-океанических рифтов за последние 10
миллионов лет. По ним можно судить об относительной скорости движения в
различных океанах.
Рис. 25. Главные плиты коры движутся, как показано
стрелками, удаляясь от рифтовых зон (черные
полосы), в которых происходит поднятие лавы, и по направлению к зонам
(заштрихованные полосы), где происходит медленное погружение коревого
вещества в глубь тела Земли. Ширина черных полос показывает ширину зон земной
коры, образовавшихся за последние 10 миллионов лет. Общая площадь этих зон
составляет 5% площади поверхности Земли. За это время такие же по площади
участки коры подверглись разрушению, погрузившись в недра Земли (в областях,
показанных штриховкой). Проекция карты выбрана таким образом, чтобы сохранить
масштаб по возможности одинаковым. Посмотрев
еще раз на рисунок 22, мы увидим множество линий, направленных с востока на
запад, по которым Атлантический рифт подвергся смещению. Эти отчетливые,
глубоко проникающие разрывы вызваны напряжениями, возникающими в коре под
действием противоположно направленных движений. Они представляют собой одну
из разновидностей геологических образований, называемых разломами. Трение при
движении блоков коры по разломам является причиной многочисленных
землетрясений. Система
желобов и горных цепей. Таким образом, ясно, что части земной коры движутся.
Движутся не только сами континенты, но и целые плиты земной коры толщиной от
50 до 100 километров, на спине" которых плавают континенты. Можно
сравнить плиту коры, несущую на себе частично погруженный в нее континент, с
широкой глыбой льда, в которую вмерзло бревно и которая плавает в озере
вместе с другими льдинами. Но
куда движутся континенты? Если материал коры создается одна полоса за другой,
в области срединно-океанических рифтов, то что-то, видимо, должно происходить
с корой, чтобы размеры ее не увеличивались и соответствовали размерам Земли.
Где-то она должна разрушаться с такой же скоростью, с какой она создается. Участки
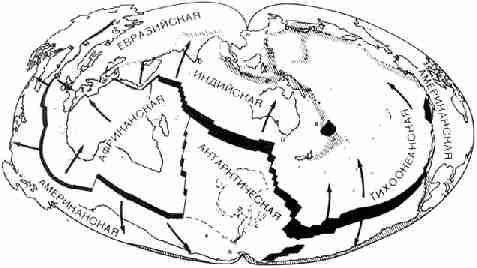
ее разрушения не трудно найти. Вокруг всего земного шара прослеживается
система узких длинных полос, показанных штриховкой на рисунке 25. Они тянутся
от Антарктиды к мысу Горн, вдоль западных окраин обеих Америк, через
Алеутские острова, Японию, Филиппины, Новую Гвинею и Новую Зеландию. Ветвь,
отходящая от этой системы, тянется на запад, через Индонезию, Гималаи, Иран,
Турцию и область Средиземноморья. Эта
огромная система состоит из трех элементов: 1) глубоководные желобы на дне океана, достигающие 11 километров глубины -
величина, превосходящая высоту самых высоких гор; 2) дугообразные ряды
вулканических островов, подобные ожерелью; 3) длинные цепи высочайших гор
Земли. Цепь, которую образуют все три элемента, представляет собой зону
нестабильности. Нестабильность указанной зоны убедительно доказывает тот
факт, что почти все самые сильные и самые глубокие землетрясения происходят в
данной зоне. Наиболее значительными землетрясениями последнего времени были
землетрясения в Чили, Калифорнии, Аляске, Иране и Турции. В
целом эта гигантская цепь дополняет собой океанический рифт огромной
протяженности. С обеих сторон к ней движутся плиты коры. Они изгибаются вниз
и поглощаются Землей и в глубине переплавляются (рис. 26), подобно восковой
свече, которую держат горизонтально и медленно вводят в горячее пламя.
Разрушение в результате плавления на одном конце происходит с такой же
скоростью, с какой образуется кора на другом конце. Поэтому общая площадь
коры остается постоянной. Процессы образования коры приурочены к океаническим
рифтам, а процессы ее разрушения - к системе желобов и гор. На обширных
площадях между этими зонами процессы происходят не так активно и состоят
главным образом в эродировании пород, накоплении
осадков и постепенном поднятии или прогибании
земной коры.
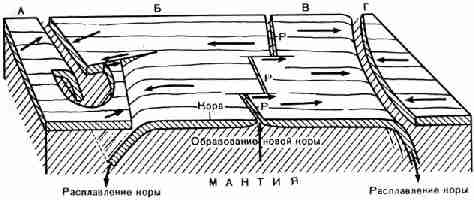
Рис. 26. Схема, показывающая образование и
разрушение земной коры и столкновения плит коры. Видны участки четырех плит (А,
Б, В, Г); стрелками показано направление их движения. Океанический рифт (Р) осложнен разломами. В краевой части плиты А на модели
прорезано отверстие, через которое видно, что в передней части плита Б
погружается под плиту А, а в задней части соотношение обратное. В другом
случае столкновения плита В погружается под плиту Г. Столкновение плит
сопровождается землетрясениями и горообразованием. Масштаб схемы не выдержан.
Длина соответствующего ей участка земной коры может достигать 5000 км.
Континенты на схеме не показаны (Р. Ф. Флинт излагает тектоническую теорию
плит. Эта теория пользуется большим распространением, но не является
единственной. - Прим. ред.) Геометрия
плит коры. Мы сравнивали плиты коры, часть из которых показана на рисунке 25,
с отдельными частями головоломки. Сравнение не слишком близкое, так как части
головоломки сохраняют свои контуры, а плиты коры надстраиваются с
"хвостовой стороны" и разрушаются путем расплавления с переднего
края. Поэтому их форма не остается постоянной. Некоторые из них сейчас,
возможно, представляют лишь осколки существовавших ранее, а другие (как,
например, Евразия) могут состоять из нескольких более древних плит, спаянных
воедино. Аналогия с конвейерной лентой хотя и полезная, но тоже не очень
близкая, поскольку плиты поворачиваются. Например, различная ширина черных
полос на рисунке 25 показывает, что в течение последних 10 миллионов лет
Тихоокеанская плита повернулась по часовой стрелке. В этом отношении плиты в
большей степени напоминают огромные льдины, которые движутся по течению в
озере, постоянно соприкасаясь и сталкиваясь друг с другом, и медленно
поворачиваясь, и даже тая по краям, и снова замерзая. Очень вероятно, что
цепи высочайших гор Земли являются результатами столкновений между плитами
коры. Например, система Альп, обнаруживающая признаки сильного изгибания,
смятия и выжимания пластов, некогда горизонтальных, могла быть образована при
столкновении движущейся к северу Африканской плиты с Евразийской. Таким же
образом Гималаи могли образоваться при столкновении Индийской плиты с
Евразийской. Гималаи все еще активны, поднимаясь со скоростью около 30
сантиметров в столетие. Слои осадочных пород, типичных для многих
континентальных шельфов (рис. 16), не могли бы выдержать, окажись они
зажатыми между двумя огромными массами, каждая толщиной в десятки километров,
сближающимися хотя и медленно, но с огромной инерцией. Маленькая лодка,
зажатая между двумя тяжело груженными баржами, скорее всего будет раздавлена
до неузнаваемости, хотя бы баржи и двигались со скоростью менее 6-8
километров в час. Когда
мы говорим о баржах, плавающих в воде, мы должны помнить, что мы описываем не
континенты, свободно плавающие и движущиеся в базальтах. Мы говорим о толстых
плитах базальта, которые движутся в каком-то еще более тяжелом веществе,
залегающем ниже, и несут на себе континенты. Поэтому по линии желобов и
хребтов сталкиваются не только континенты, но и целые толстые плиты коры с
покровом глубоководных осадков, описанных в пятой главе. Некоторые горы (как,
например, Альпы) включают большие массивы метаморфических пород, химический
состав которых указывает на принадлежность их к глубоководным океаническим
отложениям, подвергшимся сжатию. Эти осадки были слишком легкими, чтобы
погружаться, и вероятно, нагромождались вдоль желоба, по мере того как
базальтовая плита, подстилавшая их, погружалась в глубины Земли. Подобно
пене, плавающей на поверхности воды, эти отложения продолжали плавать и в
результате подверглись такому интенсивному сжатию, что были превращены в
метаморфические породы. С
помощью магнитных данных, которые мы описали раньше и которые показывают
положение магнитных полюсов в прошлом, мы можем проследить, возвращаясь в
прошлое время, вероятное положение континентов до 100 миллионов лет назад, то
есть до мелового времени. Тогда континенты находились ближе друг к другу, чем
сейчас, и Атлантический океан был гораздо уже. Попытки проследить движение
дальше в глубь времен (до ранней юры или еще более раннего времени) дали нам
гораздо менее четкие результаты. Одна из карт мира, восстанавливающая
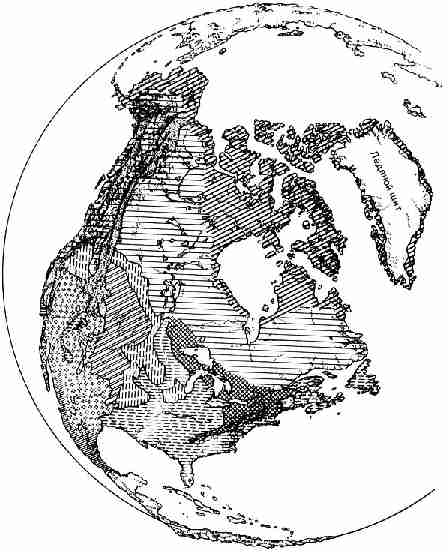
ситуацию, которая могла бы быть 200 миллионов лет назад (рис. 27), показывает
современные континенты объединенными в единый "мировой континент",
который затем распался на куски, как подтаявший весной лед на озере. На рисунке
показаны предполагаемые пути движения отдельных кусков, по которым они
достигли своего современного положения.
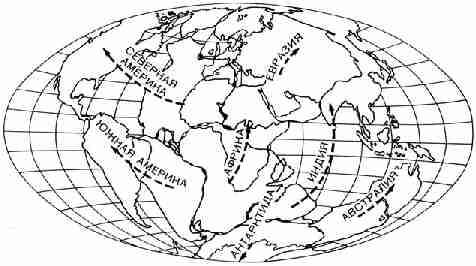
Рис. 27. Карта, иллюстрирующая пока еще не
доказанное положение о том, что около 200 миллионов лет назад (в начале
юрского периода) на Земле существовал огромный единый континентальный массив,
который затем распался на плиты. Последние перемещались в направлениях,
указанных стрелками, и создали существующие ныне континенты. Подбор
соответствующих друг другу фрагментов проводился с помощью ЭВМ Какой
бы интересной и многообещающей ни была эта карта, нужно отдавать себе отчет в
том, что она пока целиком принадлежит к области теоретических построений.
Однако сами плиты, возможно, гораздо старше 200 миллионов лет. Действительно,
они могли впервые образоваться уже 2,5 миллиарда лет назад. Если это так, то
период их существования охватывал больше половины всей истории Земли. Что
заставляет их двигаться? Читатель не может представить себе этот подвижный
ряд рифтов, желобов, движущихся плит коры и горных цепей, не задаваясь
вопросом, что же заставляет их двигаться? Каков механизм этого движения и
откуда берутся огромные запасы энергии, управляющие им? Имея представление о
системе в целом, мы не знаем, что заставляет ее работать. Несомненно, энергия
ее имеет своим источником внутреннее тепло, которое поступает на поверхность
интенсивнее всего в области океанических рифтов. Представляется вероятным,
что большая часть этого тепла создается радиоактивным распадом во внешней
части тела Земли. Но
если даже внутреннее тепло действительно дает необходимую энергию, каким
образом эта энергия приводит в движение плиты коры? Было предложено несколько
объяснений. Одно объяснение, получившее широкую поддержку, - это механизм
конвекции, процесс, который можно наблюдать в кастрюле с очень густым супом,
нагретым до кипения. Нагреваясь у дна кастрюли, жидкость медленно
поднимается, образуя беспорядочно размещенные вертикальные токи или ячейки.
Достигая поверхности, она охлаждается, растекается и опускается в других
местах, освобождая место для поднимающихся более горячих масс. Предполагается,
что конвекция происходит в мантии Земли. В простейшем виде эта идея
заключается в следующем: медленно поднимаясь в области океанических рифтов,
тяжелое вещество, обладающее свойствами жидкости, достигает коры и
растекается под ней. Удаляясь от рифтов, оно увлекает с собой расположенные
выше плиты коры. Области под желобами и горными цепями соответствуют
опускающимся звеньям конвекционной системы. Текучее вещество мантии,
несколько охлажденное, опускается, уравновешивая восходящий поток нагретого
вещества в области рифтов. Представление
об этом механизме является чисто гипотетическим и вызывает серьезные
возражения. Предлагались и другие, менее простые объяснения. Но нам придется
подождать, пока возникнет более совершенная идея, которая удовлетворит
большинство ученых. Теперь,
когда мы представили себе движение плит коры, мы видим более ясно, каким
образом гранитные породы континентов отделяются от более тяжелых базальтовых
пород. Плиты коры, состоящие из базальтов с включенными в них более легкими
материковыми породами, опускаются по системам желобов. Погрузившись на
достаточно большую глубину, они частично расплавляются, образуя более легкую
магму, чем чисто базальтовая. Эта магма, естественно, стремится двигаться к
поверхности, и образовавшиеся породы в конце концов подвергаются химической
сортировке процессами выветривания. Таким образом, движение плит способствует
тому, что с течением времени континентальная кора делается легче и
приближается по своему составу к гранитам. Заключение Мы
заканчиваем обзор наших представлений о том, как образовалась планета Земля,
как создались континенты и необыкновенная система активных рифтов, движущихся
плит и горных цепей. Мы проследили, хотя и бегло, каким образом Земля пришла
к своему современному состоянию физического тела, на поверхности которого
протекают столь разнообразные процессы. Следующий наш шаг - понять, каким
образом поверхность Земли оказалась населенной огромным количеством
разнообразных живых организмов. Глава седьмая. Молодость биосферы Органическое
вещество Предположительно
мы уже знаем, что атмосфера образовалась в результате выделения некоторых
первичных элементов из глубин Земли, где они первоначально входили в состав
ее твердого тела, и что гидросфера образовалась в результате конденсации
атмосферной влаги. Атмосфера
представляет собой простую смесь азота, кислорода, углекислого газа, водяного
пара и некоторых других газов. Состав гидросферы еще более прост - главным
образом Н2О. По сравнению с ними
биосфера гораздо сложнее. Несмотря на то что она состоит из тех же самых
основных веществ, что атмосфера и гидросфера, биосфера сложна благодаря
многочисленным химическим и структурным изменениям. Раньше, когда знания об
органическом веществе были еще очень ограниченными, основное органическое
вещество называли протоплазмой. Химическими элементами, которые образуют это
вещество (а в действительности большую группу веществ), являются главным
образом водород, кислород, азот и углерод. В самом деле, около 99% всего
органического вещества состоит из указанных четырех элементов. Вместе с ними
в состав протоплазмы входят сера, фосфор и в небольших количествах еще свыше
двух десятков элементов. Эти составные элементы образуют огромное количество
соединений. В количественном отношении суммарный состав биосферы близок к
составу морской воды. В нем не содержится ни одного элемента, который был бы
свойствен исключительно живому веществу, ни одного, который не встречался бы
в неорганической материи. Поэтому в основе химическое строение вещества
биосферы не отличается от строения двух других подвижных оболочек (гидросферы
и атмосферы. - Ред). Оно отличается только по
способу отбора и организации основных компонентов. Это различие хорошо
согласуется с идеей, которую поддерживает большинство ученых и согласно
которой живое вещество было построено из неорганической материи. Клетки Живое
вещество состоит из структурных и функциональных единиц, называемых клетками;
они могут быть уподоблены кирпичам или блокам, из которых строится здание, и,
различаясь по форме и размерам, могут образовывать множество разнообразных
функциональных систем. Каждая клетка окружена клеточной мембраной,
полупроницаемой молекулярной "стенкой", через которую могут
проходить вода и питательные вещества. Некоторые простые организмы состоят из
одной клетки; другие представляют собой чрезвычайно сложные системы клеток,
выполняющих множество разнообразных функций. Вне зависимости от степени
сложности каждый организм сохраняет свою структуру и форму, в то время как
через него проходит поток вещества, имеющего своим источником Землю, и
энергии, происходящей от Солнца. Поток этот принимает форму химических
реакций, осуществляющих разнообразные функции: питание, построение материала
новых клеток, удаление отходов и воспроизводство особей. Другими словами,
организм, созданный из клеток, живет. Период жизни отдельной клетки
колеблется от нескольких минут до нескольких десятилетий. В конце концов она
разрушается. Но тип клеток продолжает существовать, обладая способностью к
воспроизводству. Три
главные группы живых организмов Все
живые организмы объединяются в три большие группы, две из которых - растения
и животные - хорошо всем известны. В общих чертах мы можем определить
различия между ними, сказав, что животные обладают способностью к
произвольному перемещению, а растения не обладают. Однако более существенно
биохимическое различие этих групп. Растения создают свою пищу путем
химической реакции восстановления в процессе, называемом фотосинтезом.
Процесс этот заключается в том, что он воздействует на углекислый газ (СО2), отделяя кислород от углерода и используя
воду и солнечную энергию. Растения образуют органические соединения, в
которых солнечная энергия накапливается как потенциальная энергия, а
остаточный кислород выделяется в свободном виде (О2).
Животные отличаются от растений. Они питаются веществами, которые уже созданы
другими организмами, и поедают растения или других животных, которые в свою
очередь питались растениями. При дыхании они поглощают свободный кислород из
атмосферы или гидросферы и с его помощью окисляют органические соединения
пищи. Растения тоже дышат, но менее активно. Окисление
- это процесс медленного сгорания, которое высвобождает энергию из
углеродно-водородных соединений органических молекул. Это та энергия, которая
была накоплена растениями в их тканях. Окисление создает возможность
использования этой энергии во многих процессах. В процессе дыхания выдыхается
СО2, который может сразу же снова
вовлекаться в жизненный цикл растений. Таким образом, между растениями и
животными существует двусторонняя связь, которая осуществляется путем реакций
окисления и восстановления, причем основным предметом обмена служит кислород.
В реакции восстановления растения выделяют кислород; в окислительных реакциях
животные выделяют углекислый газ. Третья
группа организмов гораздо менее известна. Она охватывает царство
одноклеточных организмов, не являющихся ни животными, ни растениями; сюда
относятся зеленые водоросли, бактерии и одноклеточные простейшие, которые
прежде считались животными. Тела у большинства простейших состоят из одной
клетки или слабо дифференцированы. Предполагается, что зеленые водоросли были
предками растений. В действительности простейшие как группа в целом
предположительно являются общими предками и растений и животных, и потому
тесно связаны с проблемой возникновения жизни вообще. Давайте
вернемся на минуту к вопросу циклов, упомянутому выше. Циклически
повторяющееся движение элементов в биосфере подобно тому, которое происходит
в неорганических оболочках. В главе второй мы описали круговорот пород как серию
повторяющихся преобразований, при которых одни и те же химические элементы
меняются местами и образуют различные соединения. Позднее они высвобождаются
и образуют другие соединения, бесконечно повторяясь. Круговорот углерода и
кислорода, при котором эти элементы временно входят в состав тел растений и
затем животных, возвращаясь потом скова в растения, представляет собой
биологический цикл. Этот цикл, как и круговорот горных пород, является
составной частью движения вещества на Земле. Определение
живого вещества Как
можно определить живое вещество? Пожалуй, ближе всего к этому определению
стоит следующее утверждение: "Живое вещество состоит из компонентов,
которые могут воспроизводить себя, изменяться и воспроизводить эти
изменения". Эти способности обеспечивают передачу энергии и передачу
информации. Отметим, что сама способность к воспроизводству является
выдающимся свойством организмов. Неорганическое вещество к этому не способно.
Мы могли бы добавить, не усложняя определение, а лишь отмечая интересный
факт, что в общем молекулы органического вещества имеют большие размеры и
более сложны, чем неорганические, и поэтому обладают большей способностью к
изменениям. А постоянные изменения являлись отличительным свойством жизни на
Земле в течение всей ее долгой истории и продолжают происходить и в наши дни. Древнейшие
ископаемые организмы Поскольку
нас интересует древнейшая история биосферы, наиболее разумным представляется
начать с обзора данных о жизни до начала фанерозоя,
которые были получены при изучении древнейших ископаемых. Хотя
эти ископаемые немногочисленны и относятся к далеко отстоящим один от другого
моментам времени, тем не менее они дают достоверные сведения. Они все
датированы, хотя бы приблизительно, калий-аргоновым методом и могут быть положены
в основу летописи древнейшей истории живых организмов. Самые
древние ископаемые, найденные в горных породах (табл. 2), представляют собой
морские организмы: простейшие (бактерии) (фото 8) и растения, представленные
одноклеточными сине-зелеными водорослями. Остатки были найдены в Южной Африке
в породах, возраст которых насчитывает по крайней мере 3,2 миллиона лет. Эта
дата означает, что в момент, когда возраст Земли составлял лишь одну треть
современного, море уже было населено живыми организмами, хотя и самыми
простейшими. Это был значительный шаг вперед.
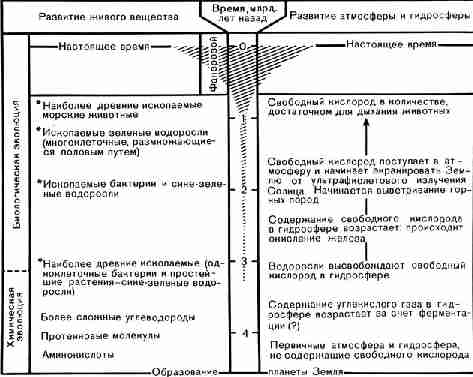
Таблица 2. Основные события истории живого вещества
и развития атмосферы и гидросферы. Заштрихованная площадь представляет
относительное количество видов организмов. Звездочками обозначены ископаемые,
возраст которых установлен калий-аргоновым методом. (Составлено по различным
источникам.) Следующие
по возрасту ископаемые остатки были найдены в Онтарио в породах,
насчитывающих два миллиарда лет. Они также представляли собой бактерии и
сине-зеленые водоросли, как показано в таблице 2. Но в породах центральной
Австралии, возраст которых предположительно около одного миллиарда лет,
найдены ископаемые растения уже нескольких видов. Среди них, кроме простых
одноклеточных организмов имеются также древнейшие из известных многоклеточных
организмов. Там же найдены зеленые водоросли - тип растений, обладающих сложными
клетками и допускающих уже половое размножение в отличие от простейшего
воспроизводства. Эти свойства допускают генетическое разнообразие и тем самым
открывают путь к развитию бесконечной дифференциации организмов. Такое
явление очевидно, возникло не более миллиарда лет назад. Но к кембрийскому
времени, в начале фанерозоя, приблизительно на 400
миллионов лет позднее, уже началось резкое увеличение количества и
разнообразия животных организмов, процесс, который привел органический мир
Земли к современному состоянию. Это "взрывное" возрастание могло
быть результатом возникновения полового размножения.
Фото 8. Древнейший из известных ископаемых организмов,
возраст которого определяется 3,2 миллиардами лет. Бактерия (называемая Eobacterium isolation) из
восточной части Южной Африки длиной менее 1/1000 мм. Фотография сделана с
помощью электронного микроскопа До
сих пор мы упоминали только простейших и растения. А что мы знаем о появлении
животных? В докембрийских слоях Южной Австралии,
вероятно насчитывающих более 800 миллионов лет, были найдены остатки сотен
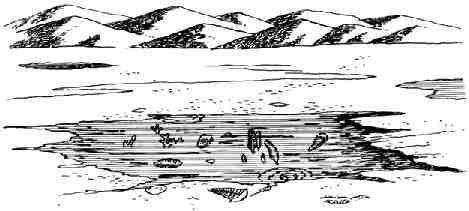
примитивных морских животных. Все они представляют собой отпечатки на мягких
отложениях древней илистой прибрежной равнины и пляжа животных, выброшенных в
отлив на берег (фото 9, рис. 28). Животные имели мягкие тела без раковин или
каких-либо твердых частей. Сохранившиеся в песчаниках отпечатки принадлежат
животным типа медуз, каким-то ветвящимся формам, напоминающим полипы, и еще
каким-то формам, не похожим ни на одно живущее сейчас животное. Наконец, были
найдены также отпечатки червей и следы их движения на мелководье. В этом
разнообразном сообществе -наиболее важным элементом являются именно черви,
так как они представляют собой вид высокоразвитых кольчатых червей. Их
наличие позволяет предполагать, что более простые животные существовали в
море гораздо раньше - настолько раньше, что они успели развиться от
простейших форм до более сложных, представленных кольчатыми червями.
Фото 9. Древнейшее ископаемое животное, найденное в
Южной Австралии - Колокольчатый червь, Spriggina Floundersi диаметром около 5 см.
Фото 9. Древнейшее ископаемое животное, найденное в
Южной Австралии - медузообразное животное, Cyclomedusa davidi, длиной около
2,5 см Едва
ли удивительно то, что мы не находим простейших животных, на существование
которых указывают сложные формы, в более древних породах. Маленькие
мягкотелые животные могли сохраниться лишь при сочетании условий, которое
встречается исключительно редко: очень тонкозернистые отложения, спокойный,
хорошо защищенный водоем, позднее заполненный осадками без размыва, и,
наконец, гораздо позднее, эрозионные процессы, которые вывели эти отложения
на поверхность, но не разрушили их. Вероятно, что большая часть еще более
древних ископаемых была уничтожена и уцелевшие становятся нам известны лишь
благодаря счастливому случаю. Существование
нескольких видов животных, найденных в Австралии, показывает, что в это время
дыхание организмов стало уже обычным процессом в биосфере. В свою очередь
наличие дыхания предполагает, что в гидросфере, а следовательно, и в
атмосфере тогда уже содержался свободный кислород, элемент, отсутствие
которого было столь характерным на первых этапах существования Земли. Как мы
увидим, источником кислорода мог быть фотосинтез растений, которые
существовали, по данным палеонтологии, к этому времени уже в течение двух
миллиардов лет или более.
Рис. 28. Реконструкция облика группы древнейших
животных, обитавших на мелководье у берегов Австралии Хотя
эти данные скудны, они все же дают нам надежную информацию не только о видах
организмов, но также косвенным образом и об изменениях в составе вод океана.
Имея в виду эти сведения, можно представить себе состав вод океана на более
ранних этапах и реакции, которые протекали в нем в начале и на ранних этапах
существования жизни. Теория
происхождения биосферы В
начале этой главы мы говорили, что живое вещество не содержит никаких
элементов, которые отсутствовали бы в неорганическом мире Земли. Добавим к
этому утверждению, что четыре главных элемента (водород, кислород, азот,
углерод), входящие в состав живого вещества, преобладают на Земле. Поэтому
живое вещество обычно. Каким
образом эти элементы объединялись, чтобы образовать биосферу? Этого мы еще не
знаем. Но мы уже имеем достаточное количество информации, чтобы сузить круг возможностей
и делать все более обоснованные предположения. Приводящий в изумление
"набор" различных организмов, живущих сейчас, включает всевозможные
градации от наиболее высокоорганизованных животных до простейших
одноклеточных форм. Но на этом последовательность не кончается, а
продолжается дальше вниз до одиночных крупных белковых молекул,
представленных, например, различными вирусами, которые существуют то в
инертной кристаллической форме, то в подвижном живом состоянии. Но
и сама белковая молекула состоит из более простых частей - соединений
углерода, водорода, азота и кислорода, связанных между собой определенным
образом и образующих так называемые аминокислоты. Эти кислоты входят в состав
любого организма, но можно ли назвать их самих "живыми"? Ученым
удалось образовать аминокислоты из соединений, которые предположительно
входили в состав первичной атмосферы Земли, - газообразной смеси водорода (Н2), метана (СН4),
аммиака (NH3) и водяного пара (Н2О). Эту смесь подвергали сильным электрическим
разрядам, а затем конденсировали. В полученной жидкости были обнаружены
аминокислоты и другие углеводородные кислоты. В других экспериментах из
неорганических составных частей были получены многие компоненты живой
материи. Результаты
этих экспериментов приводят нас к выводу о том, что организмы развились путем
длительной химической эволюции из неорганического вещества, состоявшего из
сочетаний химических элементов и неорганических соединений. Эти соединения,
наиболее вероятно, находились в морской воде, которую биологи называют
"теплым питательным бульоном", хотя это должен быть довольно жидкий
суп. Но так или иначе морская вода была жидкой средой, в которой легче всего
могли происходить химические реакции. Вода является растворителем и превосходной
средой для реакций органических соединений, которые и сами представляют собой
жидкие системы. Более того, атомы углерода обладают исключительной
способностью соединяться друг с другом самыми различными способами. Поэтому
все эти обстоятельства обеспечивали наиболее благоприятные условия для
возникновения в высшей степени разнообразных соединений. Как
мы отмечали в шестой главе, первичная атмосфера и гидросфера (включая
упоминавшийся "питательный бульон") не содержали свободного
кислорода. Весь кислород существовал только в форме соединений с другими
элементами. Кислород
не только необходим для жизнедеятельности животных; в верхних слоях
теперешней атмосферы он создает экран, защищающий Землю от ультрафиолетового
излучения Солнца, которое в противном случае убило бы все живые организмы (В
атмосфере Земли под действием ультрафиолетового излучения Солнца молекулы
кислорода (02) превращаются в молекулы озона (О3).
Озон, скапливаясь в верхних слоях атмосферы, образует экран, защищающий Землю
от ультрафиолетовых лучей. - Прим. ред.). Сейчас, мы живем как бы под
зонтиком, который образует свободный кислород, а на ранних этапах истории
Земли этой защиты не существовало. Поэтому в безжизненном первобытном мире,
даже если бы каким-то образом возник какой-нибудь современный организм, он не
мог бы выжить, потому что тогда не было этого защитного экрана. Прежде
чем двигаться дальше, следует более внимательно изучить шкалу времени,
которой нам предстоит пользоваться. Как мы уже говорили (см. табл. 2),
ископаемые остатки, найденные в породах, показывают, что приблизительно 3,2
миллиарда лет назад в море уже существовали как простейшие, так и очень
примитивные растения. Так как литосфера образовалась около 4,5 миллиарда лет
назад, мы имеем промежуток времени продолжительностью около 1,3 миллиарда
лет, в течение которого образовалось и развивалось живое вещество. Этот
промежуток времени вдвое длиннее всего фанерозоя,
со всей его долгой и сложной историей развития биосферы, что само по себе
более чем достаточно. В течение этих 1,3 миллиарда лет в среде, лишенной
свободного кислорода, должно было возникнуть и затем развиться до уровня
простейших и примитивных растений живое вещество. Рассматривая эти события,
мы должны также учитывать, что позднее кислород появился в атмосфере в
больших количествах, потому что без этого не могли бы возникнуть животные. Эти
события, последовательность которых может быть названа химической эволюцией,
теоретически могут быть объяснены логично, но насколько истинна эта теория -
покажет будущее. Компоненты,
необходимые для образования живого вещества, присутствовали в первичном
океане, этом органическом "бульоне". Без наличия свободного
кислорода ультрафиолетовое излучение Солнца свободно достигало поверхности
суши и океана. Это облучение или разряды молний могли создать высокую
энергию, необходимую для образования аминокислот, подобно тому, как это
наблюдалось в лабораторных экспериментах. Аминокислоты могли объединяться,
образуя белковые молекулы, которые, реагируя и легко вступая в соединения, способствовали
дальнейшим изменениям, создавая более сложные углеводороды. Где-то
в этой последовательности находится звено, с которого мы могли бы считать
начало возникновения жизни. Но какой момент считать началом жизни, зависит от
того, как определить жизнь. Возможно, первый живой организм состоял из одной
клетки. Возможно, это была очень простая бактерия или что-то напоминающее
бактерию. Но что бы это ни было, оно нуждалось в питании. Источником питания
для него могли служить молекулы углеводородов, составлявшие ил на дне мелкого
моря. От этого способа питания оставался один шаг до усвоения или
"поедания" сначала продуктов жизнедеятельности, а затем и мертвых
тел таких же организмов. Следующим шагом логично должно было явиться поедание
живого органического вещества. В
этой предполагаемой последовательности событий мы видим начало раздела живых
организмов на две группы. Одна группа живет, поглощая отходы
жизнедеятельности и мертвое вещество других организмов; другая группа, в
некотором отношении более сложная, предпочитает питание живыми организмами.
Это основное различие, положившее начало отдельным линиям развития, могло
привести к разделению на растения и животных, хотя эта идея, видимо,
чрезмерно упрощена. Изменения,
имевшие место в ходе этой и других ранних стадий эволюции, лучше всего
выражаются в терминах биохимических реакций. Среди наиболее ранних изменений,
вероятно, было одно, которое сделало возможной реакцию, получившую название
ферментации. В том виде, как мы наблюдаем ее сейчас, ферментация осуществляется
примитивными организмами, в том числе некоторыми бактериями, живущими в бескислородной среде. Поскольку эти организмы живут в
такой среде, они дают нам ключ к пониманию жизни, которую вели организмы в
первичном океане. При ферментации углеводороды расщепляются и
перестраиваются, небольшое количество энергии высвобождается в виде тепла, а
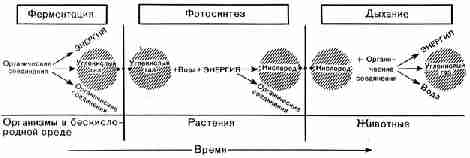
одним из образующихся продуктов является углекислый газ (рис. 29).
Рис. 29. Теоретическая последовательность процессов,
которая могла привести к образованию основных групп ныне живущих организмов.
Количество энергии, высвобождающейся при дыхании, значительно больше того,
которое высвобождается при ферментации, что показано различным размером
надписей Этот
продукт, безразлично, был ли он результатом ферментации или других химических
реакций, теоретически имел большое значение для дальнейшего развития вновь
образовавшейся жизни, потому что добавление большого объема углекислого газа
к тому его небольшому количеству, которое уже могло содержаться в море,
должно было придавать среде новые свойства. Оно создавало возможность для
процесса фотосинтеза. Свойственный многим растениям, этот процесс создает
различные органические соединения из воды, углекислого газа и энергии
солнечных лучей, поглощенной растениями (рис. 29). Если углекислый газ
постоянно поступал в море, растения, которые ранее не могли существовать в
океане, обладавшем менее сложным химическим составом, могли ^теперь
развиваться, "строя" свои ткани из углекислого газа. Отсюда
вытекает еще одно важное следствие. Одним из продуктов фотосинтеза является
свободный кислород (О2). Поэтому при
достаточном количестве растений, производящих кислород, он тоже должен был в
свою очередь накапливаться в море. В этих новых условиях в присутствии
свободного кислорода становится возможным процесс дыхания (рис. 29). Дыхание
- процесс, обратный фотосинтезу, - не только производит углекислый газ,
необходимый для растений, но также высвобождает большое количество энергии - прибль штельно в тридцать пять
раз больше, чем ферментация. Эта энергия имеет такую форму, которая может
использоваться для роста и движения организмов. При этих очевидных
преимуществах дыхание должно было сделать возможным существование животных.
Животные с пользой употребили избыток энергии, образующейся при дыхании. Они
научились свободно перемещаться, некоторые очень быстро, в поисках пищи.
Движение требовало координации частей тела, точного контроля и способности
быстро принимать сложные решения. Для этого нужен был мозг, еще одно имеющее
большую ценность отличие животных от растений. Таким
образом, возникновение биосферы начинается с химических процессов, которые
позднее приобретают характер биохимических. Последовательность этих процессов
в схематизированном виде представляется в следующем виде: аминокислоты
-> белковые молекулы -> более сложные углеводороды -> ферментация
-> фотосинтез -> дыхание Такова
химически обоснованная последовательность событий в образовании жизни в самом
простейшем виде. Она выглядит логичной, но нельзя забывать, что это все еше не более чем гипотеза. Зарождение процессов жизни
могло происходить и несколько отличным от описанного путем. Большинство
ученых считает, что причиной возникновения этих процессов был длинный ряд
случайных химических соединений, чисто случайных событий, таких, например,
как столкновение молекул в "питательном бульоне". Слово
"случайный" не должно наводить нас на мысль о том, что вся эта
последовательность событий была чисто "случайной", выражение, под
которым обычно имеется в виду "маловероятный" или даже "почти
невероятный". "Случайность" здесь означает только
"вероятность". Когда мы говорим о случайности события (такого, как
соударение молекул), мы имеем дело со статистической закономерностью. Каждое
случайное событие, каким бы маловероятным оно ни было, если рассматривать его
изолированно, становится более и более вероятным по мере увеличения числа
проб. Если вероятность события составляет 1 из 1000 в одном эксперименте, то
при 10 000 экспериментов вероятность того, что оно произойдет хотя бы
однажды, будет равной 19 999 из 20 000; такая вероятность обычно уже не
считается "случайностью", это почти уверенность. Теперь
посмотрим, сколько раз за данный промежуток времени могло произойти 10 000
попыток? Мы рассматриваем промежуток времени длиной около 1,3 миллиарда лет,
с возникновения Земли до появления растений, плюс 2 миллиарда лет или около
того до момента, о котором известно, что животные уже существовали. Сколько
раз могли произойти 10 000 попыток для каждого из таких неупорядоченных
событий за период протяженностью 3,2 миллиарда лет? Воображение человека
заходит в тупик при попытке вычислить такое огромное число. Ни один человек,
знакомый со статистикой, не отвергнет возможность возникновения химических
соединений просто на том основании, что для них могло не хватить времени.
Времени было более чем достаточно. В
какой-то момент в ходе изменений независимо от того, происходили они так, как
мы описали, или иначе, химическая эволюция ранних этапов стала биологической.
Произошло это с того момента, как организмы начали воспроизводить себя.
Граница между двумя эволюциями показана (произвольно) в таблице 2 на уровне
древнейших известных ископаемых организмов, хотя ее настоящее место должно
быть несколько ниже. Биологическая
эволюция, предположительно между одним и двумя миллиардами лет назад,
получила новое развитие благодаря возникновению полового размножения. В связи
с этим резко возросли возможности приспособления организмов к среде, как мы
увидим в восьмой главе. Обзор Настало
время снова вернуться к таблице 2, которая содержит в сжатом виде большую
часть того, о чем мы говорили в данной главе. Мы видим в таблице, что, пока существовала
первичная атмосфера, в море, в этом "органическом питательном
бульоне", накапливались аминокислоты, молекулы белков и более сложные
углеводороды. В какой-то более поздний момент начались такие процессы, как
ферментация, благодаря которой в океан поступало добавочное количество СО2. Среди простых одноклеточных растений
(сине-зеленых водорослей, которые обнаружены в ископаемом состоянии в Африке)
уже действовал процесс фотосинтеза, поставлявший в океан свободный кислород.
Помимо ископаемых растений, это доказывается и другими свидетельствами.
Осадочные пласты, имеющие возраст от 2 до 3 миллиардов лет, повсюду содержат
единичные слои окислов железа, осаждавшихся на мелководном морском дне в
результате соединения железа и кислорода в морской воде. Более молодые пласты
лишены этого вида осадочного железа, но содержат красные окислы железа,
образующиеся, как и сейчас, при окислении атмосферным кислородом железа при
химическом выветривании пород. Таким образом, представляется вероятным, что
по мере того, как строение растений становилось все более сложным, они
начинали производить путем фотосинтеза больше кислорода, чем его мог
поглотить океан. Лишний кислород начал поступать из моря в атмосферу, и
началось окисление пород в процессе химического выветривания. Одновременно
кислород, накапливавшийся в атмосфере, начал образовывать экран, защитивший
земную поверхность от ультрафиолетового излучения (Автор имеет в виду озон -
трехатомный кислород. - Прим. ред.). Когда количество свободного кислорода
стало достаточно большим, экран мог стать вполне надежной защитой. Растения,
которые ранее были вынуждены жить в глубинах океана, лишенных света, смогли
распространяться по всей водной поверхности, как это наблюдается и в наше
время. Кроме того, обилие свободного кислорода обусловило дыхание в широких
масштабах и таким образом предположительно способствовало эволюции морских
животных. Надежная защита от летальной радиации и наличие свободного
кислорода, доступного для дыхания, подготовили путь для следующих двух больших
шагов. Первым было образование и распространение планктона, огромного
множества микроскопических растений и животных, населяющих и сейчас
поверхностные воды открытого океана и служащих пищей более крупным морским
организмам. Вторым шагом, который стал возможен в это время, был выход живых
организмов из моря нa сушу. Вероятно, перемещение
происходило таким образом: некоторые растения и животные, в большом
количестве населявшие прибрежные воды, переместились в зону, затопляемую
приливом и обсыхающую при отливе. Когда они приспособились к существованию в
этой зоне, следующим логичным шагом был решительный выход на берег. Таким
образом, морские растения и животные сделали сушу местом своего постоянного
пребывания. Как
мы уже отмечали, сведения о наличии ископаемых остатков и их возрасте,
помещенные в таблице 2, представляют собой установленные факты. Другие данные
в этой таблице справедливы с различной степенью вероятности. Но
даже в том виде, в каком таблица существует, она дает нам общее представление
о том, как могло бы происходить развитие живой материи до начала фанерозоя. При этом следует помнить, что описанный выше
период развития Земли представляет собой не только начало ее истории. Он
охватывает почти всю историю Земли, занимая почти около 87% геологического
времени. Таким образом, все оставшееся время этой истории составляет всего
лишь 13% продолжительности существования Земли. Причина такого несоответствия
заключается в обилии фактических данных о жизни в течение фанерозоя.
Вследствие большого количества ископаемых остатков, относящихся к фанерозою, многообразие жизненных форм в эту эпоху
подтверждается многими интересными деталями, легко поддающимися расшифровке.
Но более старые породы подверглись полному метаморфизму, при этом большинство
ископаемых остатков, которые некогда в них содержались, оказались
разрушенными. Другие слои древних пород, которые могли содержать ископаемые
остатки, были перекрыты позднее осадочными слоями и, таким образом,
недоступны для исследования. Поэтому нам до сих пор неизвестна большая часть
фактов, принадлежавших к первому периоду истории Земли, который составляет
87% ее общей продолжительности. Таким образом, мы располагаем лишь
отрывочными знаниями, пробелы в которых приходится заполнять предположениями.
Мы надеемся, что эти предположения постепенно можно будет заменить фактически
обоснованными положениями. Нужно
иметь в виду еще одно важное положение, относящееся к описанной выше истории
Земли. Существует
не только влияние окружающей среды (какой, например, является и бескислородная атмосфера) на биосферу, но и обратное
влияние биосферы на окружающую среду. Убедительным примером такого влияния
может служить то обстоятельство, что свободный кислород, которым мы
пользуемся, был, скорее всего, внесен организмами, буквально
"выдохнут" растениями в процессе фотосинтеза. Таким образом, мы,
как и животные, обязаны царству растений не только пищей, которую мы едим,
но, очевидно, и самим нашим существованием. Без атмосферного кислорода
человек не мог бы появиться на Земле. Глава восьмая. Эволюция живых организмов Сложная
цепь Идея
образования и расплавления земной коры и движения континентальных плит,
сопровождающего эти процессы, поражает воображение как огромными размерами и
весом движущихся глыб, так и колоссальной энергией, которая необходима для
поддержания всей этой системы в движении. Однако не менее поразителен процесс
эволюции живых организмов, проходящий через всю историю планеты Земля; он
вызывает изумление огромным количеством участвующих в нем организмов и
крайней сложностью биохимических процессов, в совокупности образующих Жизнь.
В течение всей исгории Земли отдельные растения и
животные, умирая, захоронялись под отложениями, а
их форма и строение "консервировались" в виде ископаемых остатков,
включенных в пласты пород. В наши дни многие ископаемые были найдены и
собраны воедино. Если расположить их по порядку, то видно, что они образуют
непрерывные ряды. Некоторые изменения, прослеживающиеся в поколениях
современных организмов, представляют собой непосредственное продолжение цепи
изменений, установленных по ископаемым остаткам. Все вместе эти ряды образуют
сложную цепь со многими звеньями, которая постоянно удлиняется уже более 3
миллиардов лет, с тех самых пор, как химическая эволюция была замещена самовоспроизводством организмов. Теория этой цепи, теория
эволюции живых организмов, позволяет понять историю жизни, ранее
представлявшуюся беспорядочным скоплением ископаемых форм. А кроме того, как
мы уже отметили в главе третьей, эта теория сделала возможной корреляцию
слоев в различных районах и таким образом способствовала построению
геохронологической шкалы. Когда
мы делаем попытку обрисовать историю организмов, мы закономерно начинаем с
рассмотрения процесса эволюции, который восстанавливаем по ископаемым
остаткам. Мы начнем с данных, на которых основано наше понимание этого
процесса, а затем перейдем к рассмотрению того, как работает его механизм. Доказательства
эволюции За
последние сто лет среди ученых стало господствующим убеждение, что эволюция
действовала всегда и действует и в настоящее время. Это убеждение основано на
огромном количестве фактов, в том числе на данных различных отраслей науки. В
частности, оно основано на анализе соотношения различных ископаемых и
соотношения между зародышами различных животных, анатомических данных, опыте
целенаправленного разведения растений и животных. Ископаемые.
Пожалуй, наиболее убедительным обоснованием теории эволюции являются
собранные воедино сведения об ископаемых остатках. При сравнении ископаемых,
собранных из различных пластов, становится очевидно, что чем моложе породы,
тем более сложные организмы в них обнаруживаются; усложнение происходит в
направлении от основания к вершине геохронологической колонки. Это
подтверждается на примере слоев, выходящих в Большом каньоне Колорадо. Как мы
отмечали в седьмой главе, наиболее древние ископаемые остатки принадлежат
одноклеточным организмам, следующие по возрасту - растения, а затем -
наиболее простые животные. Это прогрессирующее усложнение прослеживается к
вышележащим (то есть более молодым) слоям, и наконец, последним в позднекайнозойских слоях появляется человек. Едва ли
будет ошибкой считать это соотношение доказательством того, что со времени
начала эволюционного процесса биологическая эволюция непрерывно создавала новые
формы и структуры. Если
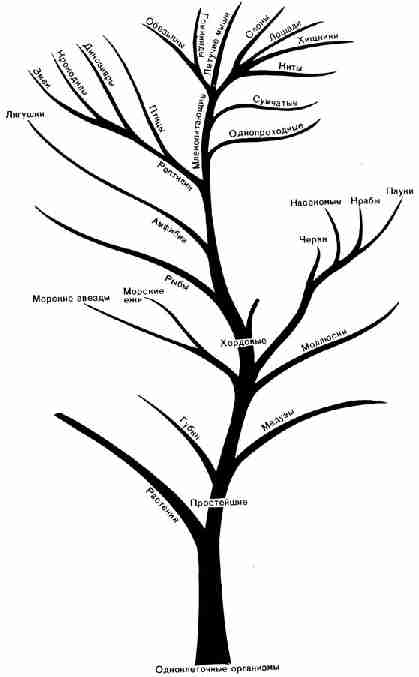
сравнивать различные ископаемые остатки не с их "родными" слоями, а
между собой и с другими организмами, которые живут сейчас, выявляется другая
удивительная закономерность. Если объединить между собой наиболее близкие
группы, то графически соотношение между ними может быть представлено в виде
дерева, со стволом, большими и малыми ветвями, причем существующие сейчас
виды помещаются на вершинах ветвей (рис. 30). Если проследить распределение
этих организмов по ветвям, сверху вниз, изменения кажутся незаметными, но
общий эффект при переходе от малых ветвей к большим может оказаться весьма
значительным. При переходе от малых ветвей, к большим ископаемые организмы
становятся все более простыми, причем самые простые располагаются у основания
ствола. Так или иначе, но все эти жизненные формы связаны друг с другом, как
имена предков в родословном древе. Даже крошечные простейшие являются
предками человека, как мы можем установить, проследив эту линию достаточно
далеко вниз.
Рис. 30. "Древо жизни", показывающее
соотношение между различными формами жизни, как современными, так и
ископаемыми. Приведенная схема далеко не полная; в нее включены только
наиболее известные группы животных и не дано подразделений растительного мира Конечно,
"древо жизни" до сих пор еще не "укомплектовано", и хотя
это и не показано на рисунке 30, но в некоторых случаях связи между отдельными
ветвями еще не прослежены. Пробелы в ряду ископаемых остатков представляют
собой недостающие звенья в цепи наших данных. Но постепенно одно за другим
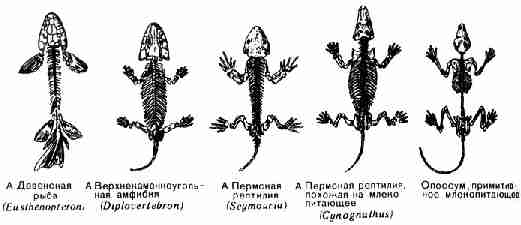
недостающие звенья находятся и занимают свое место. Таким звеном явилась,
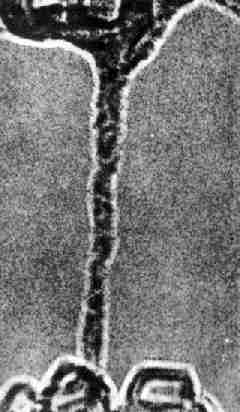
например, одна из древнейших и наиболее примитивных ископаемых амфибий - Ichthyost$ga, найденная в девонских слоях в Гренландии в
1948 г. Она настолько похожа на девонских рыб, что только конечности
указывают на принадлежность ее к наземным животным (рис. 31). Это указывает
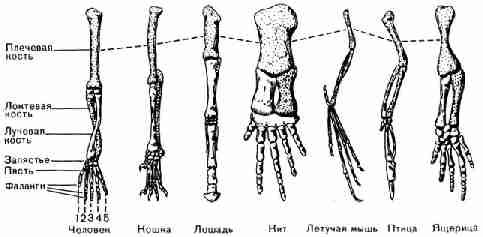
на связь между рыбами и амфибиями.
Рис. 31. Скелеты рыбы и семи наземных животных; с
помощью одинакового по размеру изображения скелетов, данных в одном и том же
положении, показано сходство основных частей и строения скелетов Другое
некогда недостающее звено - археоптерикс (Archeopterух),
самая примитивная из известных нам птиц, впервые обнаруженная в юрских
пластах в Германии в 1861 г. Эта "покрытая перьями рептилия"
представляет собой промежуточное звено между рептилиями и птицами и отнесена
к птицам потому, что она покрыта перьями. Наконец, древнейший человек -
австралопитек (Australopithecus, africanus) (фото 64), обнаруженный в Южной Африке в 1924
г., значительно более примитивный, чем известные нам до сих пор по ископаемым
остаткам древние люди. Он образовал новое звено в цепи примитивных предков
человека.
Рис. 31. Скелеты рыбы и семи наземных животных; с
помощью одинакового по размеру изображения скелетов, данных в одном и том же
положении, показано сходство основных частей и строения скелетов Некоторые
ветви эволюционной цепи оказались гораздо длиннее, чем это предполагалось
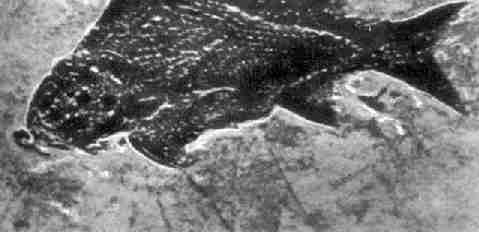
раньше. Например, в 1938 г. рыбаки в Индийском океане поймали очень большую
странного облика рыбу, целаканта (фото 10). Эта
примитивная рыба со своеобразными лопастевидными
плавниками была хорошо известна в ископаемом состоянии и встречалась в
пластах начиная от девонского и до мелового возраста, однако считалась
вымершей еще до начала кайнозоя. Открытие живого целаканта
продлило историю существования этого вида еще на 70 миллионов лет. Эти рыбы -
близкие родственники девонских рыб с лопастевидными
плавниками, из которых развились амфибии, как мы уже отмечали выше, - также
образуют звено в цепи предков человека.
Фото 10. Целакант,
"живое ископаемое". Экземпляр, пойманный в марте 1966 г Другим
"живым ископаемым" является разновидность секвойи, Metasequoia (фото 11), которая до 1941 г. считалась
исчезнувшей в среднекайнозойское время. В 1941 г.,
однако, она была открыта в одном из внутренних районов Китая. Сейчас эти
деревья, родственные хорошо знакомой нам секвойе, выращиваются из привезенных
из Китая семян и в других странах.
Фото 11. Метасеквойя,
выросшая в Соединенных Штатах из семени, присланного из Китая Сходство
зародышей. Другие доказательства эволюции были получены при изучении
зародышей. Развивающиеся зародыши различных видов позвоночных обнаруживают
поразительное сходство между собой на ранних стадиях своего развития, но по
мере Дальнейшего развития утрачивают это сходство. Чем ближе родство между
видами животных (как, например, между человеком и обезьяной), тем дольше
сохраняется сходство между их развивающимися зародышами. Особо
следует отметить наличие одинаковых жаберных щелей, которые видны на ранних
стадиях развития у различных видов животных. Эти щели, однако, изменяются у
различных животных по мере развития. У рыб жаберные щели и связанные с ними
органы образуют приспособление для дыхания жабрами. Но птицам и
млекопитающим, которые дышат иначе, жаберные щели не нужны для дыхания. Они
преобразовались в органы, с помощью которых животные издают или слышат звуки.
У человека, например, из эмбриональных жаберных щелей развилась часть уха и
горла. Это является достаточно веским доказательством того, что человек и
другие дышащие воздухом позвоночные сохранили эмбриональные черты,
унаследованные от далеких предков, которые дышали жабрами и жили по крайней
мере 360 миллионов лет назад. Сходство
скелетов. Если сравнивать скелеты древних и современных наземных позвоночных
(рис. 31), они свидетельствуют приблизительно о том же, о чем и зародыши.
Скелеты состоят из сходного количества костей, располагающихся приблизительно
в одном и том же порядке. Только размеры отдельных костей и их относительные
пропорции меняются у различных животных. Это сходство, столь устойчиво
прослеживающееся, может означать только одно: скелет современных наземных
позвоночных в основе своей представляет очень старомодную конструкцию. Хотя с
течением времени его отдельные части удлинялись или укорачивались,
увеличивались или уменьшались, но в целом он никогда не был заменен чем-то
совершенно новым. Он только постоянно, и то очень медленно, преобразовывался
путем многочисленных, но очень небольших изменений. Таково еще одно
неопровержимое доказательство того, что современные высшие животные развились
из своих отдаленных предков, многое от них унаследовав. Еще
более ясно это можно увидеть, если мы подробно рассмотрим какие-либо части
скелета, например переднюю конечность (рис. 32). У всех позвоночных передняя
конечность имеет одинаковое в основном строение, но у каждого вида она
изменилась таким образом, чтобы обеспечить наибольшее удобство при
передвижении в данных условиях среды обитания - в воздухе (птицы), в море
(киты), на обширных травянистых равнинах (лошади) и в лесу (кошки). Таким
образом, как мы вскоре увидим, среда оставляет свой отпечаток на скелетах.
Рис 32. Кости передней конечности различных
позвоночных. Строение в целом у всех одинаково, но в различной степени
видоизменено и приспособлено к определенному типу передвижения: полету
(летучая мышь, птица), плаванию (кит), быстрому бегу (лошадь), прыжкам
(кошка). У ящерицы и человека несколько отличны основные черты строения Рудиментарные
органы. Как кости, так и мягкие части тел многих животных организмов образуют
органы, которые не несут очевидных функций, но сходны с органами других
живущих или вымерших организмов, у которых эти органы выполняли вполне
определенные функции. У тех животных, у которых эти органы бесполезны, они
носят название рудиментарных, потому что являются остатками - рудиментами -
приспособлений, существовавших у предков. Только в теле человека таких
органов более 150; наиболее известен из них аппендикс, слепой отросток,
который может воспаляться и во многих случаях подлежит хирургическому
удалению. У некоторых менее развитых животных, однако, соответствующий орган
функционирует как полезная часть пищеварительного тракта. Менее известным
органом является копчик. Он состоит из семи позвонков с соответствующими
мускулами и нервами и расположен у основания позвоночника. Это рудиментарное
образование, которое в более развитом виде образует хвост у большинства
животных. Малые берцовые кости, у современных лошадей не выполняющие никаких
функций, представляют собой остатки когда-то использовавшихся пальцев у
предков лошади, имевших три опорных пальца. Киты и некоторые змеи имеют
рудиментарные кости, которые соответствуют костям задних конечностей у
четвероногих животных. Некоторые нелетающие птицы, например страус, имеют
рудиментарные крылья - остатки крыльев, которые другие птицы используют для
полета. Таковы немногие примеры, выбранные нами из огромного множества
рудиментарных органов, существование которых может быть объяснено только тем
фактом, что они стали рудиментарными в ходе эволюции. Одомашнивание
животных и растений. В результате направленного разведения в течение
длительного времени - от нескольких лет до нескольких тысяч лет - человеку
удалось вывести многочисленные разновидности отдельных видов растений и
животных. Наиболее известный пример - многочисленные породы собак, очевидно,
выведенные от одного первоначального вида, напоминавшего волка. Породы собак
очень разнообразны по размерам, форме тела и соответствию специальным целям,
хотя все они принадлежат к одному виду. Эти искусственно полученные изменения
показывают возможность передачи по наследству признаков, отобранных
человеком, и позволяют предполагать, что подобный процесс изменений имел
место и в течение длительной истории организмов. Процесс
эволюции После
обзора наиболее убедительных доказательств происходившей эволюции, мы можем
обратиться к рассмотрению самого процесса, в котором выделяются три аспекта.
Первый - молекулы, которые могут копировать сами себя и передавать в
закодированном виде информацию. Второй - изменения, которые происходят с
отдельными организмами и могут быть унаследованы. Третий - среда и ее влияние
на состав популяций. Если внимательно рассмотреть эти три аспекта, то
нетрудно будет понять, как именно происходила и происходит эволюция, зафиксированная
по ископаемым остаткам. Молекулы
ДНК. В любом организме каждая отдельная клетка, имеющая ядро, содержит
некоторое количество очень важного вещества, называемого
дезоксирибонуклеиновой кислотой (сокращенно ДНК). Молекула ДНК имеет
необычные для единичной молекулы большие размеры, и все же, чтобы сделать с
нее фотографию обычного размера, она должна быть увеличена в сотни тысяч раз.
Фотография, сделанная с помощью электронного микроскопа, показывает, что она
имеет форму двух переплетенных спиралей (фото 12). Большая спиральная
молекула ДНК обладает замечательной способностью к самовоспроизводству.
Она может делиться на две совершенно равные половины, каждая из которых
содержит половину спирали. Затем каждая половина может достраивать
недостающую часть, снова приобретая вид спирали. Результатом является
удвоение - две полных спирали вместо одной, существовавшей ранее. Считается,
что это удвоение возможно потому, что каждая часть исходной спирали состоит
из четырех химических компонентов, которые могут располагаться в различном
порядке. Подобно точкам и тире азбуки Морзе, долгое время использовавшейся
при передаче телеграмм, порядок, в котором расположены химические компоненты,
составляет генетический код, который содержит информацию или "инструкции",
согласно которым должна строиться вторая половина спирали. Закодированное
таким образом послание представляет собой своего рода чертеж. Оно определяет,
какие химические элементы должны использоваться, а также в каком порядке они
должны располагаться. Порядок образует химические соединения, которые нужны
клетке или группе клеток для того, чтобы развиваться и функционировать.
Конечно, эти химические компоненты, служащие для построения клеток,
получаются из пищи, поглощенной и переработанной организмом, причем количество
пищи должно быть достаточно велико.
Фото 12. Изображение молекулы ДНК, полученное с
помощью электронного микроскопа при увеличении в 1 750 000 раз. Эта молекула
была получена из клеток тела рыбы; она образует длинную узкую вертикальную
полосу, которая для удобства фотографирования помещена между двумя
скоплениями кристаллов углерода (вверху и внизу на снимке). При данном
увеличении видна двойная спираль, но составляющие ее неразличимы. Фотография
автомобиля, сделанная при этом увеличении, имела бы размеры от Нью-Йорка до
Сан-Франциско Именно
так отдельный организм (набор специализированных клеток) растет в продолжение
всей своей жизни путем деления и перестройки клеток, согласно закодированным
инструкциям, содержащимся в молекулах самих клеток. По мере того как клетки
делятся, инструкция тоже воспроизводится, так что любая клетка, имеющая ядро,
получает свою собственную копию и может немедленно ее использовать. Таким
образом определяется не только рост, но и каждая черта индивидуального
организма. Если трудно представить себе такую большую детальность, следует
вспомнить, что молекула ДНК, какой бы большой она ни была, все же очень мала
с точки зрения человека. Диаметр ее составляет около 0,0000008 сантиметра, и
в теле человека может находиться почти неисчислимое количество таких молекул.
Сложность процессов, которые одновременно происходят в таком большом городе,
как, например, Нью-Йорк с его восьмимиллионным населением, не очень велика по
сравнению со сложностью процессов образования, движения и разрушения молекул
ДНК и других составных частей в теле одного обитателя этого города. Резюмируя,
можно сказать, что молекулы ДНК с записанной в них информацией и способностью
воспроизводить самих себя определяют основной план роста и функционирования
клеток, составляющих организм, и, следовательно, все основные черты и
особенности организма. Мы должны посмотреть, каким образом передается
закодированная информация, а следовательно, и особенности организма, от
одного поколения к другому, так как эта информация лежит в основе
наследственности. Наследственность
и изменчивость. Характеристики формы и поведения, определенные генетическим
кодом, передаются от родителей к потомству, но в этом процессе они меняются,
по крайней мере до некоторой степени благодаря строению клеток, участвующих в
процессе полового размножения. В отдельной особи любого вида организмов,
размножающихся половым путем (а сюда относятся почти все животные и
растения), половые клетки содержат длинные нитевидные хромосомы, каждая из
которых в свою очередь содержит многочисленные гены, - единицы,
контролирующие наследственность. Ген - это часть молекулы ДНК, та часть,
которая содержит в своем строении запись информации, необходимой для
воспроизводства данного организма - его формы, размера, физиологии и
поведения. И
вот когда мужская клетка спермы объединяется с женской яйцеклеткой,
образуется новая сложная клетка, имеющая комбинацию хромосом обоих родителей.
Оплодотворенная клетка начинает делиться. При этом делении точно
воспроизводится новая комбинация хромосом. Код, которым обладает эта сложная
клетка, конечно, отличается от кода любого из родителей. В соответствии с
этим и новая особь может обладать чертами родителей почти в любом сочетании.
Количество сочетаний, которые могут образовывать хромосомы в одном
человеческом организме, оценивается в 2×1010. Это почти
невероятно большое число. Поскольку потенциальное число комбинаций настолько
велико, две особи не могут быть в точности одинаковыми. Каждая из них
получает гены от обоих родителей, и эти гены образуют уникальную комбинацию.
Эта комбинация содержит неповторимый набор закодированных сведений и
инструкций, согласно которым развиваются индивидуальные черты организма, до
некоторой степени передающиеся по наследству следующим поколениям. Существенным
результатом этого является изменчивость. Поскольку каждый индивид в каждом
поколении до некоторой степени отличается от своих родителей, изменчивость
постоянно проявляется в виде многочисленных мелких, незаметных изменений,
которые могут быть унаследованы. Естественный
отбор. На этой стадии сказывается влияние окружающей среды. Каждый организм в
каждом поколении рождается со своим собственным комплексом изменений в
определенных условиях среды. Этот организм реагирует на условия,
приспосабливаясь к ним, но степень этого приспособления частично будет
зависеть от унаследованных изменений. Некоторые особи из большой популяции
будут в большей степени соответствовать условиям среды, чем другие, просто
потому, что один или несколько свойственных им признаков дали им небольшое
преимущество в этом окружении. Поэтому статистически у одной группы особей
будет чуть больше шансов выжить и воспроизвести себя, чем у остальных.
Поскольку изменения могут наследоваться, следующее поколение будет иметь
немного большее количество особей, обладающих благоприятными изменениями.
Если условия останутся неизменными, то приобретение всей популяцией
"благоприятных" свойств явится только вопросом времени, хотя,
возможно, довольно длительного. Это
настолько важный вопрос и так часто его понимают неправильно, что мы вернемся
к нему еще раз. В любой популяции изменения проявляются в каждом поколении.
Из этих постоянно возникающих наборов изменений условия среды стремятся
отбирать лучшие (то есть наиболее соответствующие условиям). В результате
этого создается впечатление, что отдельные особи в популяции сознательно
пытаются приспособиться. Однако ничто не может быть дальше от истины. Этот
процесс приспособления есть не что иное, как ряд приспособительных реакций.
Эволюция не имеет программы, но точность, с которой приспосабливающиеся
организмы отвечают на требования среды, похожа на чудо. Простой,
но вместе с тем ясный пример этого процесса представляет история эволюции
бабочек в Англии за последнее время. Если сопоставить данные наблюдений и
подсчетов, сделанных в настоящее время, с теми, которые были сделаны в 1850
г., то окажется, что около 60 видов бабочек в Англии изменили свой облик. За
этот период времени в результате развития промышленности здания и даже
деревья покрылись копотью. В 1850 г. менее 10% популяции бабочек имели темную
окраску, сейчас же темный цвет имеют более 90%. Первоначальное соотношение
цветов было результатом изменчивости. Однако так как светлая окраска была
более заметна на темном фоне, птицы легче находили и истребляли бабочек со
светлой окраской и меньшее их количество выживало и размножалось. Для бабочек
критическим фактором среды явилось наличие насекомоядных птиц. Бабочки не
приспосабливались сами сознательно к более темной окраске среды; они
оказались к ней приспособленными в результате деятельности птиц. Выживание
под влиянием среды и с помощью механизма наследственности определяет облик
грядущих поколений и называется естественным отбором. Это процесс сортировки
и перетасовки. Хотя естественный отбор действует в сфере организмов, он
подобен по своим результатам отбору в неорганическом мире, сортировке и
перемешиванию, которые производят другие природные процессы, создающие земную
кору. Глубоко под поверхностью Земли определенные минералы
"отбираются" благодаря относительно низкой точке их плавления, и их
расплав движется вверх. На поверхности минералы "отбираются" в
соответствии с их устойчивостью по отношению к процессам химического
разрушения, их неустойчивые компоненты удаляются. Действительно, естественный
отбор более понятен, если мы рассматриваем его как один из многих сложных
природных процессов, протекающих на Земле. В
естественном отборе изменение от одного поколения к другому может быть
незначительным, но, как и в случае врезания Большого каньона Колорадо или
сортировки частиц, образующихся при выветривании гранита, кумулятивный эффект
бывает огромным. Как мы отметили в предыдущих главах, на поверхности Земли
непрерывно происходят геологические изменения, вызывающие изменения природных
условий. Эти последние в свою очередь влияют на растения и животных, вызывая
адаптацию. Мы
должны помнить, что такие изменения происходят не сознательно и не целенаправленно.
Они действуют автоматически, слегка изменяя вероятность выживания каждой
особи в популяции. Каждая особь, со своим набором присущих только ей свойств,
сталкивается с требованиями среды. Некоторые особи выживают, другие
оказываются истребленными, и каждое новое поколение становится немного лучше
приспособленным к условиям среды, чем были приспособлены его родители. Повсюду
в мире мы наблюдаем растения и животных, которые в процессе приспособления приобрели
строение и черты, наиболее соответствующие данному виду среды. Белый медведь,
обладающий толстой шкурой с жировой подкладкой, прекрасно умеющий плавать,
хорошо приспособлен к жизни на полярных льдах. Длинноногий верблюд, со своей
способностью сохранять воду в организме, приспособлен к жизни в обширных
районах континентов, отличающихся малым количеством воды и скудной
растительностью. Другие организмы приспособились к жизни в тропических лесах,
в воздухе над обширными океанами, в пещерных водоемах и ко многим другим
условиям среды. Каждое из этих приспособлений явилось результатом эволюции,
направлявшейся естественным отбором. Если мы внимательно изучим ископаемые,
мы можем прийти к выводу, что в течение большей части истории организмы
приспосабливались к тем же видам природной среды, которые существуют и
сейчас. Но эти организмы меняли свое пространственное размещение вследствие
геологических изменений, затрагивавших и поверхность континентов, и океаны.
Такое приспособление организмов в прошлом явилось еще одним следствием
принципа актуализма. Мутации.
Имеется и другой вид наследуемых изменений. Он увеличивает разнообразие живых
организмов, среди которых под влиянием окружающих условий происходит отбор.
Это мутации. Они происходят в яйцеклетке или сперматозоидах одной особи и
могут наследоваться потомством. Мутации заключаются в изменении кодового
механизма внутри молекулы ДНК, так что некоторый ген у особи нового поколения
отличается от соответствующего гена у любого из родителей. Происходят они неожиданно
и непредсказуемо и, будучи наследуемыми, вызывают изменения структуры, черт
или характеристик, которые проявляются в последующих поколениях.
Взаимодействуя со средой, эти изменения закрепляются в потомстве, если они
благоприятны, или "выбраковываются", если они неблагоприятны.
Химические основы мутаций в деталях еще не изучены, но можно полагать, что
эти изменения имели существенное влияние на направление эволюционных
изменений организмов. Дискуссия Эволюция
в основе своей состоит в адаптации - приспособлении к окружающей среде.
Изменения происходят постоянно - они имеют или характер мутаций или
происходят в результате новых сочетаний свойств, возникающих в последующих
поколениях. Они поставляют сырой материал, над которым работает естественный
отбор. При этом испытываются всевозможные виды новых приспособлений, и те из
них, которые оказались "успешными" в данных условиях, сохраняются и
в будущем, неудачные же постепенно отсеиваются. Когда
Чарлз Дарвин впервые описал естественный отбор в
1859 г., он рассматривал этот процесс с точки зрения приспособленности
организма для жизни в данной среде, конкуренции между отдельными особями,
преуспевания некоторых групп и исчезновения менее приспособленных. В
рамках геологической истории успех групп организмов зависит от двух факторов:
1) способности эффективно воспроизводиться и 2) способности занимать
территорию и удерживать ее в условиях конкуренции. В истории динозавров,
цветковых растений или мамонтов ледникового периода все эти вопросы не имели
никакого отношения к морали. Мораль - это человеческое понятие, и оно
вступает в силу лишь на поздних.этапах истории
человека. Следует
добавить, что изучение летописи ископаемых остатков не приводит нас к
установлению какой-либо общей цели эволюции. Не доказано, что эволюция
следовала в каком-либо определенном направлении и стремилась к определенной
цели. Другими словами, эволюция, очевидно, не имеет программы. Очевидно, она
не следует также и по прямой линии. Ее пути часто переплетаются (как мы
видели на рис. 30) и отклоняются от прямого направления, но они никогда не
повторяются в точности. Такой путь развития мы определяем как вероятностный,
являющийся результатом непрерывного ряда ответных реакций живых организмов на
новые возможности, возникающие при изменении среды. Сколько
времени потребовалось для того, чтобы процесс эволюции создал все то огромное
разнообразие растений и животных - более миллиона видов, - которое существует
сейчас? В начале этого процесса, как ,мы отмечали в седьмой главе, постепенно
возникли первые одноклеточные организмы. Это было более 3 миллиардов лет
назад. Ближе к нашему времени органический мир достиг в основном своего
современного состояния к концу плиоцена, возможно 2-3 миллиона лет назад.
Большая часть эволюционных процессов протекла в интервале между этими двумя
моментами. Никого, кто изучал этот процесс, не удивит то обстоятельство, что
развитие организмов могло достичь современной степени приспособленности и
разнообразия за отрезок времени немногим более 3 миллиардов лет. Ученые считают,
что этого времени было вполне достаточно. Действительно, если бы
геологические процессы, которые вызывали изменения среды обитания, протекали
быстрее, то возможно, что та же степень эволюционного развития, которую мы
видим сейчас, могла быть достигнута за более короткое время. Скорость
эволюционных изменений, конечно, менялась в широких пределах в зависимости от
времени и места, была неодинаковой у различных видов организмов и зависела,
как и сейчас, от скорости и пространственного распределения изменений
окружающей среды. Например, набор ископаемых остатков показывает, что за 63
миллиона лет геологического времени, то есть за кайнозойскую эру,
беспозвоночные морские организмы развивались медленно и изменились
незначительно, а большинство наземных животных развивалось гораздо быстрее и
при этом сильно изменялось. Эта разница отражает различия в скорости
изменения среды. Природная обстановка на суше быстро реагировала на поднятия,
горообразование и изменение температур и атмосферных осадков, в то время как природные
условия в море оставались сравнительно стабильными. И все же с точки зрения
человеческого календаря любая скорость эволюции кажется малой. За период,
предположительно составляющий 10 000 лет, в течение которого собаки и скот
были одомашнены и подверглись воздействию искусственного отбора, не
образовалось ни одного нового вида. Искусственные изменения привели не более
чем к появлению разновидностей в пределах одного вида. Мы
должны подчеркнуть этот основной принцип. Пока окружающие условия остаются
стабильными, популяция организмов тоже остается стабильной, с небольшими
эволюционными изменениями. Когда же среда начинает изменяться и, таким
образом, становится неустойчивой, населяющие ее организмы также становятся
нестабильными. Таким образом, окружающая среда создает предпосылки для
проверки биологических "изобретений", которые постоянно появляются.
Когда мы проследим историю живых организмов, восстановленную по ископаемым
остаткам, мы увидим много примеров таких "изобретений"; некоторые
из них оказались успешными, а другие, не выдержав испытаний, исчезли с лица
Земли. Тесная
связь организмов со средой, их восприимчивость к изменениям среды, которая
ясно прослеживается по ископаемым остаткам, найденным в древних пластах,
должна послужить уроком современному человеку. На территориях, где в сильной
степени развита промышленность, подчас происходят резкие изменения условий
жизни. Некоторые из этих изменений более заметны и невыгодны для организмов
(по крайней мере некоторых), чем изменения, обусловленные климатическими
колебаниями или другими природными процессами. Изменения обоих видов, как
можно ожидать, вызывают соответствующие ответные реакции (приспособление или
постепенное вымирание) в биосфере этих территорий. Возможные последствия
этого не следует недооценивать человеку - единственному виду, который
вызывает изменения и может их контролировать с учетом конечных результатов -
благоприятных или катастрофических для животных и растений Земли. Глава девятая. Особенности ископаемых остатков Палеонтологическая
летопись В
нашем обсуждении геохронологической шкалы и эволюции органического мира часто
упоминалась летопись ископаемых остатков, или палеонтологическая летопись.
Под этим термином подразумевается вся совокупность собранных, определенных,
занесенных в каталоги и описанных ископаемых остатков различных видов,
которые хранятся в музеях, университетах и других научно-исследовательских
учреждениях во всем мире. Палеонтологическая летопись включает сотни
коллекций и миллионы больших и малых ископаемых, от огромных динозавров и
китов до микроскопических животных и растений. Сюда относятся не только сами
ископаемые, но и все, что о них написано, целые библиотеки научных книг и
статей, в которых содержатся описания и рисунки ископаемых и рассматривается их
положение в геохронологической шкале. К настоящему времени полностью описано
100 000 видов ископаемых морских беспозвоночных, и это число постоянно
увеличивается. Таким
образом, составление летописи ископаемых - очень большое и важное дело, в
котором участвуют тысячи ученых и технических специалистов, и это становится
очевидно для каждого, кто посетит какой-либо институт, занимающийся изучением
ископаемых остатков. Каждый любознательный посетитель хочет знать, как
выглядят ископаемые, как могут сохраняться животные и растения в течение
миллионов лет и каким образом в случае необходимости производится
реконструкция ископаемых. Ответ на эти вопросы сделал бы нашу историю живых
организмов гораздо более понятной. Что
мы могли бы знать об истории Земли, если бы организмы, населявшие Землю в
прошлом, хотя бы частично не сохранились в ископаемом состоянии? Конечно, мы
знали бы о геологических циклах, о принципе актуализма и движении плит земной
коры. Но наша шкала геологического времени была бы очень примитивной и гораздо
менее полезной, чем сейчас, наши реконструкции природных условий прошлого
гораздо менее полными, а пути эволюции можно было бы восстанавливать лишь
предположительно. Сохранение
органических остатков Существованием
ископаемых в довольно большом количестве мы обязаны процессу консервации
некогда живых организмов. Хотя большая часть отдельных организмов, некогда
живших на Земле, была уничтожена, местами их тела или части тел оказались
защищенными от воздействия кислорода. Они были быстро погребены под осадками
и затем превращены в породы в результате осаждения минерального вещества из
медленно просачивающихся вод, пропитывающих грунт. Такова в общих чертах
история большинства ископаемых. Давайте посмотрим более внимательно, каким
путем сохраняются органические остатки. Когда
животное или растение умирает, большая часть его вещества довольно быстро
подвергается разрушению. Другие организмы, среди которых особенно важную роль
играют бактерии, пожирают или химически разлагают его. Обычной реакцией в химическом
разложении как органического, так и неорганического вещества является
окисление. Мы видим в лесу, что ствол упавшего дерева, пролежавший на земле
несколько лет, становится бурым и сгнивает. Ископаемые остатки, встречающиеся
в пластах пород, составляют лишь ничтожную часть когда-то живших организмов,
избежавшую общей участи. В виде исключения их тела были защищены от окисления
и трупоядных животных, что обычно происходило
благодаря погружению в воду в момент смерти или вскоре после него. Огромное
большинство ископаемых остатков принадлежит морским беспозвоночным -
различным моллюскам и им подобным организмам. Мягкие ткани их тел или заживо
съедались хищниками, или пожирались животными-могильщиками после смерти этих
организмов. Но твердые раковины сохранялись и заносились осадками,
отлагавшимися на дне моря. На суше возможность сохранения была гораздо
меньше. Но даже там некоторые животные и растения попадали в живом или
мертвом виде в воду, которая предохраняла их тела от окисления. Другие быстро
погребались под накапливающимися осадками, которые вскоре после этого
насыщались грунтовыми водами. Часто захоронение как животных, так и растений
происходило при паводках на реках (фото 13). Другой причиной сохранности
могло быть быстрое накопление вулканического пепла, а иногда и текущей лавы.
Короче говоря, главными факторами, обеспечивающими сохранность остатков на
первых этапах, были быстрое погребение или погружение в воду, или и то и
другое вместе. Почти такое же значение имеет третий фактор - наличие в
мертвом организме твердого вещества (раковины, костей, зубов или рогов),
которое противостоит действию трупоядных организмов
и химическому разложению. Почти все ископаемые остатки состоят из одних
скелетов; мягкие ткани сохраняются редко и только в совершенно особых
условиях. Однако, если тело захороняется целиком,
оно может сохраняться до мельчайших деталей (фото 4). Даже игра красок на
крыльях ископаемых насекомых сохранилась в течение сотен миллионов лет.
Фото 13. Это песчаное русло временного водотока
большую часть времени сухое или почти сухое. При сильных ливнях по нему течет
мутный поток, в котором тонут животные, а их трупы уносятся течением и быстро
погребаются в аллювии Способы
сохранения. Образование ископаемых остатков не обязательно заканчивается
после захоронения. Они могут подвергаться и другим воздействиям, в том числе
таких процессов, которые полностью меняют их состав или разрушают вещество,
сохраняя внешнюю форму. Наиболее
простой пример представляет тело, которое оказалось заключенным в лед,
асфальт или другое природное вещество, которое сохраняет материал этого тела.
Именно так случилось с 36 мамонтами и несколькими шерстистыми носорогами,
найденными в северной Сибири за последние 200 лет; их трупы сохранились в
грунтах, находившихся в мерзлом состоянии со времени последнего оледенения.
Мясо, по крайней мере одного из них, было настолько свежим, что его ели
ездовые собаки. Распространено ошибочное мнение о том, что некоторые из этих
мамонтов были найдены во льду ледников. Это не так. Насколько нам известно,
они были заключены в промерзшем иле. Возможно, животные утонули в болотах как
раз перед осенними заморозками, при этом ил промерз на большую глубину, и на
следующий год, ранней весной, еще до того, как он оттаял, речное половодье
отложило толстый слой аллювия. Этот слой препятствовал таянию залегавших ниже
пород. Радиоуглеродные определения возраста, сделанные для шкуры и шерсти
этих животных, показали, что между самым ранним и самым поздним захоронением
прошло более 30 000 лет. Таким образом, если взять только 36 найденных
мамонтов, то в среднем захоронение животных, утонувших в трясине, должно было
происходить только раз в 800 лет. По сравнению с этим число мамонтов, которые
погибли более обычным способом, должно быть гораздо больше. Согласно
подсчетам, из Сибири были вывезены в виде слоновой кости бивни 20 000-30 000
мамонтов, найденных на поверхности. Возможно, еще большее количество бивней и
целых туш все еще погребено в мерзлом иле. Другой
пример - это туши шерстистых носорогов ледниковой эпохи, сохранившиеся даже с
характерной для них длинной шерстью в нефти, которая сочилась из земли и
затем затвердевала. В восемнадцатой главе описан случай захоронения костей
миллионов млекопитающих в нефтяном озере в Калифорнии (такие же захоронения
есть и около города Баку. - Ред.). Химизм сохранения остатков в нефти во
многом сходен с мумификацией человеческих останков, использовавшейся древними
египтянами. Бальзамировщики использовали нефть, которая не пропускает воздуха
и потому препятствует окислению.
Фото 14. Окаменелое дерево. Срез небольшого ствола.
Клеточная структура сохранилась вследствие заполнения клеток кремнеземом Гораздо
чаще - практически всегда - мягкие части трупов разрушаются, а твердые
подвергаются окаменению. Этот процесс, в основе химический, протекает в толще
грунта и представляет собой часть медленно протекающего процесса превращения
осадков в осадочные породы. Медленно просачивающаяся грунтовая вода отлагает
минеральные вещества в пористых внутренних частях костей или в полостях
клеток растений (фото 14). Грунтовая вода может также растворять первичное
вещество раковин или костей и замещать его каким-либо другим веществом,
обычно кремнеземом, иногда в виде опала; реже, при соответствующем химизме
среды, происходит замещение каким-либо редким минералом, как, например,
чистым серебром. Наконец, окаменение может совершаться путем естественной
возгонки, происходящей глубоко под землей. При этом процессе водород и
кислород, содержавшиеся в исходном органическом веществе, медленно выделяются
в виде различных газов и остается только углерод. Таким образом, форма
организма, очень часто это бывают рыбы, сохраняется в виде почти чистого угля
(фото 15).
Фото 15. Рыба длиной 15 см (род Redfieldius),
сохранившаяся в углефицированном виде в результате
естественной возгонки в триасовых сланцах (Дурхэм Конн, США) Хотя
большая часть известных нам ископаемых представляет собой окаменелые раковины
и кости, существуют и другие способы консервации организмов. В слоях ила и
глины, отложенных на дне глубоких озер, часто встречаются отпечатки листьев
(фото 16). Листья опускаются на дно озера и постепенно покрываются
отложениями. Вещество листа разлагается, но отпечаток его формы остается.
Иногда таким же образом сохраняются отпечатки насекомых (фото 36).
Фото 16. Отпечаток листа платана в эоценовых
алевролитах, штат Юта. Сам лист разрушен; темный цвет объясняется наличием
минерального вещества, отложенного грунтовыми водами во время превращения
первичного осадка в горную породу Редкая
разновидность отпечатка была обнаружена в слое базальта - лавовом потоке
олигоценового или миоценового возраста - около города Каули,
штат Вашингтон. Двадцать-двадцать пять миллионов лет назад на пути этого
потока, наступавшего на мелководное озеро, оказался лежавший на боку труп
носорога длиной более двух метров; труп был заключен в лаву, которая в воде
остыла настолько быстро, что не разрушила органических тканей. Таким же
образом лава залила и росшие поблизости деревья. Открытые
в 1935 г. остатки носорога представляли собой небольшую полость в теле
лавового потока, имевшую форму раздувшегося трупа носорога (рис. 33); в этой
полости находились челюсть, зубы и другие кости животного. Для изучения формы
тела носорога был сделан гипсовый слепок полости.
Рис. 33. Реконструкция позы трупа ископаемого
носорога (Вашингтон). (Пояснение дано в тексте.) Еще
один вид ископаемых остатков, никогда не содержавших органического вещества,
- это отпечатки следов (фото 17). Из них сохраняются только те следы, которые
были перекрыты тонкими отложениями, прежде чем их успела разрушить эрозия.
Иногда даже крохотные следы мелких насекомых (фото 18), а также отпечатки
дождевых капель сохраняются очень отчетливо. Такие отпечатки не просто
редкость. Они содержат информацию о размере животного, длине конечностей,
наличии когтей, копыт и т.д., а также о том, как передвигалось это животное -
шагами, бегом иди прыжками. Другой, на первый взгляд странный, но надежный
источник сведений - это окаменевшие экскременты животных. Изучение их тонких
срезов под микроскопом помогает определить, какую пищу употребляли животные,
и неопровержимо свидетельствует о том, что некоторые из них были каннибалами.
Они питались животными того же вида.
Фото 17. Цепочки следов, в большинстве длиной 25-45
см, оставленных динозаврами на поверхности напластования триасовых
песчаников. Государственный заповедник динозавров, Коннектикут. Более 200
миллионов лет назад этот песчаник был песчаным аллювием. Стрелками указаны
гораздо более мелкие следы, возможно, принадлежащие животному, изображенному
на рис. 47 Таковы
основные способы образования и консервации ископаемых остатков - глубокое
промерзание тел, окаменение костей и раковин, естественные отливки, отпечатки
следов; всем им сопутствует захоронение тем или иным способом, обычно очень
быстрое (фото 19).
Фото 18. Следы насекомых и птиц в алевролитах,
оставленные около 45 миллионов лет назад на побережье эоценового озера в
штате Юта Реконструкция.
Для того чтобы ископаемые остатки могли по-настоящему изучать специалисты,
они должны быть реконструированы. Некоторые ископаемые, особенно морские
беспозвоночные (двустворчатые моллюски, улитки, устрицы), почти не требуют
реконструкции, кроме разве подбора соответствующих друг другу створок
раковины. Но реконструкция ископаемого позвоночного животного - задача
гораздо более сложная. Чаще всего отдельные кости оказываются сдвинутыми или
разбросанными. Прежде чем собирать их, каждую кость нумеруют (или помечают
как-то иначе) и тщательно фотографируют в том положении, в каком она была
найдена. Затем, уже в лаборатории, они очищаются от вмещающей породы и
подбираются по порядку специалистом, знающим анатомию.
Фото 19. Палеонтологи откапывают кости огромного
динозавра диплодока в юрских песчаниках во впадине Бигхорн,
Вайоминг Восстановление
скелета вымершего животного основывается на общих анатомических
закономерностях, но реконструкция мягких тканей, кожи и шерсти не может быть
совершенно точной, потому что она может осуществляться только по аналогии с
ныне живущими родственными видами. Хотя о различных, менее очевидных
анатомических подробностях можно только догадываться, все же ископаемые
остатки рассказывают нам гораздо больше, чем это кажется с первого взгляда.
Остатки наземных животных включают не только кости, зубы, рога и пластинки
панциря, но также превосходные отпечатки шкуры и следы, сохранившиеся до
мельчайших подробностей. В некоторых случаях можно судить об умственных
способностях животного по форме и величине гипсовой отливки внутричерепной
полости, содержавшей мозг. Конфигурация черепа позволяет также судить об
органах чувств - зрения, слуха, обоняния - и предположить, могло ли это
животное издавать звуки. Иногда ископаемые кости сохраняют даже следы
болезней животного. При восстановлении облика вымерших млекопитающих, живших
во время последнего оледенения, современным ученым приходят на помощь
художники каменного века, которые оставили на стенах пещер изображения
животных (на которых они охотились) в том виде, в каком они их наблюдали
(рис. 34).
Рис. 34. Точное изображение вымерших млекопитающих
ледниковой эпохи, сделанное художниками позднего каменного века. Вверху
шерстистый носорог. Внизу найденное в Западной Франции изображение
нападающего мамонта, вырезанное на куске клыка 10 000-16 000 лет назад.
Художник не вырезал до конца клыки, хобот и передние ноги Классификация Вид.
Классификация живых существ берет начало в весьма отдаленных временах, когда
считалось, что каждая разновидность была создана отдельно. Позже, когда был
понят процесс эволюции, классификация стала еще более необходимой, так как к
этому времени уже было открыто множество организмов, которые должны были быть
систематизированы в целях изучения, и число их непрерывно возрастало. Без
систематической классификации ископаемые были бы бесполезны, как большая
библиотека без каталога. Основная
единица в классификации - вид, группа организмов, которые могут скрещиваться
между собой, но не с особями из других групп и обладают общими для целой
группы особенностями формы и строения. У ныне живущих организмов в основу
выделения вида может быть положена возможность к взаимному оплодотворению.
Для вымерших организмов, к которым относится большинство известных ископаемых,
виды могут устанавливаться только по аналогии с существующими группами
организмов или по особенностям формы и строения. Вид
образуется только потому, что члены этого вида каким-либо образом были
изолированы в отношении размножения от соседних популяций, так что гены этого
вида не могли смешиваться с другими. Если эта изоляция продолжалась довольно
долго, то эти гены становились несовместимыми с генами соседних групп. С
этого момента изолированная группа становится видом по определению. С другой
стороны, если члены группы не были изолированы, то они могли свободно
скрещиваться с членами соседних групп, и тогда со временем отличительные
признаки этой группы должны исчезнуть. В ходе истории Земли геологические
процессы постоянно вызывали изменения условий, создавая при этом различные
типы изоляции. Поэтому вполне понятно, что почти столько же времени, сколько
существует жизнь, должны существовать и виды. Но благодаря медленным
геологическим изменениям, протекающим непрерывно, одни виды вымирают, а другие
перемещаются, изолируются и дают начало новым видам. Подтверждение
этого процесса было найдено в меловых породах, слагающих обрывы английского
побережья, обращенные к проливу Ла-Манш. Мел - это белая осадочная порода,
состоящая главным образом из миллионов микроскопических раковин, образованных
из кальцита. Он накапливался на морском дне в течение части мелового периода.
Кроме этих крошечных раковин, мел содержит многочисленные остатки морских
ежей вида Micraster. Большое количество их было
найдено в пласте мела толщиной не менее 150 метров, который отлагался в
течение 4 миллионов лет. Морские ежи в основании пласта сильно отличаются от
найденных в верхней части, причем это изменение происходит постепенно.
Морские ежи из нижних горизонтов составляют одну группу видов, в то время как
найденные в верхней части принадлежат к совершенно другим видам, развившимся
путем эволюции из первых. Этот
постепенный переход показывает трудности проведения границ между ископаемыми
видами, для которых возможность скрещивания не может быть проверена. Поэтому,
как мы уже отмечали, ископаемые виды определяются на основе сходства формы и
строения. Каталог
Линнея. В середине XVIII в., за сто лет до того,
как идея эволюции получила общее признание, швед Карл фон Линне
(Линней), который писал по-латыни и подписывался Linnaeus,
потратил многие годы на составление каталога живых организмов. Он
классифицировал растения и животных по их очевидным анатомическим свойствам.
Как и другие ученые в это время, Линней считал разнообразные живые организмы
однажды сотворенными и затем более не изменявшимися. Данная статическая
концепция представляет в настоящее время лишь исторический интерес, но
каталог Линнея имеет все же большую научную ценность, представляя собой
первичную основу современной классификации организмов. В своей основе он не
подвергся изменениям, за исключением деталей, и кроме того, он написан
по-латыни, на этом почти универсальном языке ученых. Название каждого вида
организмов состоит в этом каталоге из двух слов. Первое слово обозначает
более широкое понятие - род, второе, более узкое - вид. Например, Homo sapiens (человек
разумный), Pinus strobus
(белая сосна), Felis domesticus
(кошка домашняя), Felis leo
(лев), Felis tigris
(тигр), Felis pardus
(леопард). Все
более крупные подразделения последовательно накладываются на категории,
использованные Линнеем. Так, два или более родственных видов образуют род,
два или более родственных родов образуют семейство, два или более семейств -
отряд, два или более отрядов - класс, два или более классов - тип. Два или
более типов составляют царство, наиболее крупную категорию, так как три
царства включают соответственно все одноклеточные организмы, растения и
животных. Современная
классификация. Для наших целей нам нет необходимости давать в этой книге
полную детальную классификацию организмов. Поэтому мы исключаем одноклеточные
организмы и даем нашу сокращенную классификацию животных и растений
соответственно в таблицах 3А и 3Б. Теперь,
когда мы точно знаем, что такое вид, естественно возникает вопрос, сколько же
существует различных видов животных и растений. По наиболее точным оценкам,
сейчас насчитывается около 1,5 миллиона живущих видов. Из них 860 000
составляют насекомые, 350 000 - растения, 8600 - птицы и только 3200 -
млекопитающие. Большая часть остальных видов, около 300 000, относится к
морским беспозвоночным. Общее количество - 1,5 миллиона - включает только те
виды, описания которых были опубликованы учеными. Считается, что вдвое
большее количество видов еще не описано. Если это так, в настоящее время
существуют 4,5 миллиона видов. В это число не входят вымершие виды, известные
только в виде ископаемых остатков. Основываясь на количестве уже описанных
ископаемых видов, общее количество вымерших видов, обитавших когда-либо на
протяжении более трех миллиардов лет существования жизни на Земле, оценивают
в пределах от 50 миллионов до 4 миллиардов. И это огромное число относится
только к видам. Только когда мы вспомним, что каждый вид может включать
множество особей, живущих одновременно (например, в 1972 г. на Земле жило 4
миллиарда людей - членов вида Homo sapiens), тогда мы начнем хотя бы в слабой степени
представлять себе размеры и сложность строения биосферы. Ископаемые
и стратиграфия Изменения
уровня моря и эволюция. Теперь мы знаем, как живые существа становятся
ископаемыми и как классифицируются ископаемые, чтобы их можно было
максимально использовать в научных целях. Располагая этими данными, мы можем
теперь сказать больше о связи ископаемых с последовательностью осадочных
пластов. Например, правомерно будет спросить, какое влияние окажут наступания и отступания моря на организмы, живущие в море
вблизи берега. Таблица
3А Царство
животных Тип
Protozoa (простейшие - одноклеточные, большей
частью микроскопические животные) Тип
Porifera (губки) Тип
Coelenterata (кишечнополостные - кораллы, морские
анемоны, медузы и др.) Тип
Brachiopoda (плеченогие) Тип
Bryozoa (мшанки) Тип
Echinodermata (иглокожие - морские звезды, морские
ежи, морские лилии и др.) Тип
Mollusca (моллюски - улитки, двустворчатые
моллюски, устрицы, кальмары, осьминоги и др.) Тип
Arthropoda (членистоногие - креветки, крабы, омары,
трилобиты, пауки, насекомые) Тип
Vertebrata (Chordata)
(позвоночные и хордовые - животные, имеющие спинную струну) Класс
(в действительности, 4 класса) Pisces (Рыбы) Класс
Amphibia (Амфибии или земноводные - лягушки, жабы,
саламандры) Класс
Reptilia (Рептилии или пресмыкающиеся - крокодилы,
ящерицы, змеи, черепахи, динозавры и др.) Класс
Aves (Птицы) Класс
Mammalia (Млекопитающие - теплокровные животные,
вскармливающие детенышей молоком) Таблица
3Б Царство
растений Несосудистые растения: Водоросли
(простейшие, преимущественно водные растения) Грибы
и плесени Мхи
(простейшие наземные растения) Сосудистые
растения: 1.
Примитивные сосудистые растения 2.
Лепидодендроны, плауны 3.
Хвощи 4.
Папоротники и семенные растения Папоротники Голосеменные
(семенные папоротники, цикадовые, гинкго, хвойные) Покрытосеменные
(Цветковые растения) Мы
допускаем, что в воде живет столько морских животных и растений, сколько
может прокормиться при данных условиях. Те особи, которым не хватает пищи,
или умирают, или переселяются. Мы должны также помнить, что пища состоит из
имеющегося органического вещества, иначе говоря, источником питания живых организмов
(Это справедливо для животных. - Прим. ред.) могут служить только другие
живущие здесь же организмы. При этом образуются длинные пищевые цепи. Чтобы
полностью использовать всю доступную пищу, организмы путем естественного
отбора приспособились ко всем изменениям своего жизненного пространства, даже
самым малым. Например, в прибрежной зоне различные виды живых существ
населяют: 1) воды различной глубины; 2) различные виды дна - скальное, чистое
песчаное, илистое; 3) участки, различающиеся по степени защищенности -
открытый берег, подверженный действию прибоя, заливы с тихой водой и т. д. Когда
прибрежные воды окажутся таким образом полностью заполненными
взаимосвязанными между собой организмами, любое изменение уровня моря
относительно суши нарушает существующее неустойчивое равновесие и вызывает
изменения в популяции. У берегов с пологим шельфом, как, например, восточный
берег Северной Америки от Массачусетса до Флориды, на протяжении около 1600
километров понижение уровня только на 6,5 метра вызовет обсыхание
полосы шириной 8-11 километров. Площадь этой зоны составит по крайней мере 13
тысяч км2. При том, что это пространство
уже заселено почти до предела, такое изменение скажется на миллиардах больших
и малых организмов. Многое будет зависеть от скорости отступания моря, но
каким бы медленным оно ни было, в конце концов жизненное пространство будет
уменьшено на 13 тысяч км2.
"Лишние" обитатели дна, те, которые в меньшей степени способны
приспосабливаться к новым неустойчивым условиям, или вымрут, или переселятся
на новое место. Но если они переселятся, то они должны будут выдержать
конкуренцию с коренными обитателями той территории, на которую они
вторгнутся. Такая
конкуренция способствует процессу эволюции. Поэтому если последовательные
изменения уровня моря у рассматриваемого нами побережья разделены достаточно
большим промежутком времени - скажем, порядка миллиона лет, - то мы вправе
ожидать, что при повышении уровня моря, создающем добавочные площади морского
дна, которые могут быть заселены, среди организмов, которые их заселят,
появятся новые виды, эволюционировавшие с тех пор, как их предки были
вытеснены из этого района. Эти новые виды добавят новые черты к комплексам
ископаемых, составляющих палеонтологическую летопись. Они, несомненно, окажут
большую помощь ученым, которые будут решать проблему соотношения пластов. Вспомнив
главу третью, мы можем заключить, что изменения, подобные только что
описанным, являются главной причиной того, почему определенные группы
ископаемых приурочены к определенным пластам, что делает возможным
сопоставление пластов в различных районах. Вымирание.
Ход процесса эволюции в течение фанерозоя
сопровождался потерями - вымиранием видов, родов и даже более крупных групп.
Одной из главных причин такого вымирания является, как мы видели, изменение
площади морского дна, вызванное колебаниями уровня моря. Вероятно, причинами
могли быть и изменения температуры и других климатических факторов, резкое
изменение количества пищи, развитие особенно хорошо приспособленных хищников. Часто
выдвигается предположение, что основной причиной вымирания видов является
изменение климата или других факторов природной среды, вызванное изменением
магнитного поля Земли; однако до сих пор это предположение не доказано
фактами. Хотя наш календарь обращений (рис. 20) охватывает только последние
несколько миллионов лет, обращения установлены также и в мезозое и палеозое,
а возможно, случались и раньше. Наибольшее количество вымфших
групп обнаруживается в конце палеозоя и в конце мезозоя. Эти два промежутка
времени отмечены сравнительно большой частотой магнитных обращений. Сейчас
исследуется вопрос о связи этих двух видов событий. Располагая
некоторыми сведениями об ископаемых, мы можем теперь перейти к обзору
организмов, населявших Землю в прошлом, чему до сих пор мы уделяли мало
внимания. В следующей главе мы расскажем, как можно восстанавливать природные
условия прошлого по данным об ископаемых остатках и пластах, в которых они
найдены. Глава десятая. Природные условия далекого прошлого Облик
Северной Америки в наши дни В
главах первой и шестой мы уже говорили о поверхности вращающейся Земли.
Сейчас мы снова рассмотрим ее, но уже не с точки зрения ее структуры или
истории, а обращая внимание прежде всего на то, как выглядит сегодня
поверхность континентов - равнины, горы, реки и многочисленные организмы,
живущие непосредственно на поверхности Земли, над ней или под ней. Иначе мы
могли бы сказать: "... и многочисленные организмы, приспособленные к
условиям поверхности континента". Эта приспособленность явилась
результатом воздействия условий среды на бесчисленные предшествовавшие данным
организмам поколения. Если пренебречь частностями, то в течение последних 50
миллионов лет большая часть Северной Америки - с Аппалачскими горами на
востоке, горной системой Кордильер на западе и широкой полосой низменностей и
равнин, заключенной между ними и принадлежащей к бассейнам реки Миссисипи и Гудзонова залива, - в общем имела тот же облик, что в
наши дни. За это время - 50 миллионов лет - организмы успели достаточно
хорошо приспособиться к окружающим условиям, а там, где произошли
значительные геологические изменения, они успели приспособиться и к этим
изменившимся условиям заново. Поэтому в настоящее время в Северной Америке
существуют тысячи вариантов условий природной среды, каждый из которых
обладает своим собственным "набором" соответствующих данным
условиям растений и животных, совместно обитающих, служащих пищей друг для
друга, а в некоторых случаях помогающих друг другу. Понятие
"окружающая среда" включает все физические, химические и
биологические аспекты рассматриваемой территории независимо от того, большая
она или маленькая, - очертания суши, глубину водоемов, химический состав и
мутность воды, течения, климат, почвы, растительность и животный мир. Факторы
природной среды столь многочисленны, что обычно мы ограничиваемся изучением
влияния одного фактора на другой или всей окружающей среды в целом на
какой-либо интересующий нас фактор. Так, например, нас может интересовать,
как влияют температура, глубина и соленость воды на организмы, живущие в
какой-либо части моря. Или, поднимаясь на высокий горный хребет, мы можем
задаться вопросом о том, как влияет возрастание высоты на растения,
произрастающие на горных склонах. В настоящее время предметом всеобщего внимания
становится влияние дыма и газов, содержащихся в воздухе, на здоровье жителей
города. Характер
распределения растительности. Распространение различных видов растительности
на континенте дает хорошее общее представление об основных условиях обитания
человека и наземных животных. На рисунке 35 показаны естественные
растительные зоны Северной Америки (без учета изменений, вызванных сельским
хозяйством). На указанном рисунке зоны отделены друг от друга отчетливыми
линиями, но в действительности переходы от одной зоны к другой постепенны.
Сразу же бросается в глаза, что на пространственное распределение
растительности сильно влияет расположение гор. На востоке узкая длинная
полоса лесов северного типа протягивается вдоль Аппалачей более чем на 950
километров к югу, чему способствует более прохладный и немного более влажный
климат в горах по сравнению с равнинами. На западе подобное распространение
особых типов растительности в виде узких полос прослеживается в Скалистых
горах и других высоких горных массивах. Последовательно рассматривая
территорию Соединенных Штатов от Аппалачей до Скалистых гор, мы видим
последовательную смену лесов прериями и далее сухими степями. Эта смена
поясов растительности обусловлена главным образом осадками, количество
которых убывает по направлению к западу. Если подобным же образом
рассматривать центральную и восточную Канаду, следуя с юга на север, можно
наблюдать смену отчетливо выраженных лесов северного типа, субарктических
лесов и арктической тундры. Существование этих поясов обусловлено главным
образом температурами и отчасти осадками; и те и другие постепенно снижаются
к северу. На арктическом побережье Канады и Аляски общее количество осадков,
выпадающих как в виде дождя, так и снега, невелико; во многих местах оно не
превышает количества осадков, выпадающих в Аризоне. Приведенные примеры
показывают, что в целом распределение типов растительности легко может быть
понято, если его рассматривать с точки зрения основных климатических
факторов.
Рис. 35. Естественные растительные зоны современной
Северной Америки. Обобщенно показаны типы растительности, которые занимали бы
территорию, если бы там не было земледелия
Условные обозначения: 1 - арктическая тундра (злаки
и травы, деревьев нет); 2 - субарктические леса (ель, пихта, сосна, береза)*;
3 - леса северного типа (сосна, ясень, береза, тсуга)*; 4 - горные хвойные
леса; 5 - леса умеренного пояса (дуб, гикори, сосна и др.); 6 - прерия и
саванна (степь с отдельными деревьями); 7 - сухие равнины с травянистой
растительностью; 8 - пустынные кустарники и злаки; 9 - субтропические
кустарники; 10 - сухие редколесья тропического типа (Имеются, по-видимому, в
виду и леса умеренного пояса. - Прим. ред.) Местные
сообщества живых организмов. При пристальном рассмотрении каждый из
упомянутых выше поясов оказывается состоящим из многих более мелких единиц.
Взглянув на какую-либо небольшую часть континента, мы сразу же видим мозаику
еще меньших участков, каждый из которых имеет свой тип растительности в
соответствии с высотой, крутизной склонов, количеством осадков и почвами. Не только
растительность, но и животный мир меняется от района к району. Рисунок 36,
хотя на нем и не изображается какая-либо реальная территория страны, дает
общее представление о возможном сочетании местных природных условий. В левой
части схемы показано морское мелководье с песчаным дном, на котором обитают
различные беспозвоночные, рыбы и морские водоросли. Характерные обитатели
пляжа - крабы, птицы и насекомые. В полосе дюн, созданных в результате
переноса ветром в глубь территории отсортированного прибоем песка, растут
жесткие злаки, обитают насекомые и, вероятно, грызуны.
Рис. 36. Схема соотношения некоторых типов природной
среды, предположительно распространенных на западном побережье Северной
Америки и прилежащей части континента Позади
пояса дюн расположены степи и кустарниковые заросли (включающие различные
виды кустарников); дифференциация их может быть обусловлена различиями почв.
По степи протекает извилистая река, берега которой окаймлены влаголюбивыми
деревьями (например, ивами); в ней живут рыбы, пресноводные моллюски, улитки
и другие более мелкие организмы. Во встречающихся здесь озерах есть рыбы,
беспозвоночные, водоросли, тростники и камыши, привлекающие водоплавающих
птиц. Из наземных животных могут водиться олени, антилопы, койоты и грызуны. Еще
дальше в глубине континента находятся горы; их склоны в нижней части покрыты
травянистой растительностью, а выше - широколиственными лесами, переходящими
еще выше в вечнозеленые леса, и вблизи вершин - альпийские луга с различными
злаками и травами. В лесах мы можем встретить оленя, лису, горного льва
(пуму), а выше - сурков и кроликов. Горы способствуют выпадению осадков, но
по отношению к территориям, расположенным с подветренной стороны от них, они
играют роль барьера; поэтому за горами находится пустыня с разреженной
растительностью, состоящей из кустарников и кустистых злаков. Среди ее
обитателей могут быть антилопы, американские жесткошерстные кролики и другие
грызуны, койоты, ящерицы, змеи и скорпионы. В соленых озерах пустыни можно
найти беспозвоночных и некоторых мелких рыб, приспособившихся к жизни в
соленой воде, а по берегам - водяных птиц. Все эти разнообразные растения и
животные приспособились к среде, в которой они обитают. Местами
мозаичность контрастирующих между собой природных условий выражена в еще
более мелком масштабе. Например, в долине склон южной экспозиции может быть
покрыт травянистой растительностью, в то время как противоположный склон,
обращенный на север, может быть лесистым; это происходит в первую очередь
потому, что на солнечной стороне влага испаряется из почвы гораздо быстрее,
чем на теневой стороне, где влага задерживается в почве, способствуя
произрастанию деревьев. Этим объясняется и различие популяций животных,
обитающих на разных склонах, в особенности насекомых и мелких грызунов. И
действительно, каждый, кто совершал неторопливое путешествие по какой-либо
местности, размышляя над тем, что он видел, мог бы перечислить несколько, а
возможно, и много, местных типов природных условий, в которых живые организмы
прекрасно приспособились ко всем изменениям условий жизни. Окружающая
среда в прошлом Только
сейчас, когда Земля перенаселена видом Homo sapiens, человек начинает познавать природную среду и
взаимосвязи ее отдельных типов. Но ученым, изучающим природу, давно известно,
какое большое значение имеет влияние, оказываемое средой на развитие всех
организмов. Они пришли к пониманию того, что условия среды определяют
пространственное распространение растений и животных, а также сложный путь их
эволюции. Принцип
актуализма гласит, что горы, равнины и плато, реки и озера, аллювиальные
днища долин, дельты, пляжи, песчаные пустыни и другие формы поверхности суши
и дна мелководного моря, которые существуют сейчас, должны были существовать
в прошлом (фото 20). Находки ископаемых свидетельствуют, что в любой момент
геологической истории растения и животные были приспособлены к существовавшим
тогда условиям в такой же степени, как их современные аналоги к существующей
среде. Это соответствие настолько ясно, что даже там, где нет прямых
свидетельств, ископаемые формы вымерших организмов считаются доказательствами
существования природных условий, в общих чертах сходных с теми, в которых
обитают наиболее близкие к ним современные организмы.
Фото 20. Волноприбойная рябь в песчаниках Дакоты,
выходящих в Скалистых горах, в Колорадо. Хотя возраст этой ряби, возможно,
насчитывает 100 миллионов лет, она не отличается от ряби, образующейся вдоль
современных побережий. Эта рябь была создана на мелководье медленным
течением, двигавшимся слева направо, в море, показанном на рис. 38. На
величину отдельных гребней указывают современные листья осины Природные
условия, восстановленные по стратиграфическим данным. Как ископаемые
организмы, так и физические свойства пластов, в которых они обнаружены,
содержат сведения о распределении гор, мелководных морей и других крупных
форм на континентах в прошлом. Правда, сами континенты медленно перемещались
по поверхности Земли вместе с несущими их плитами земной коры, причем
движение это могло происходить как непрерывно, так и прерывисто. Чем более
удален от нас момент времени, тем менее мы можем быть уверены в том, где
находился каждый континент в этот момент. Но несмотря на затруднения,
проистекающие из этого, мы можем установить положение прежних мелководных
морей, гор и т.п. в пределах любого отдельного континента, например Северной
Америки. Мы можем даже составить серию схематических карт континента,
показывающих, как менялись во времени основные черты его поверхности. Большей
частью наши исследования ограничиваются последними 600 миллионами лет или
около того, то есть отрезком времени, который в таблице 3А
обозначен как фанерозой. Как уже говорилось, более
древние слои содержат слишком мало ископаемых остатков, а их первичные
физические свойства были изменены или уничтожены метаморфизмом. Вышесказанное
приводит нас к следующему выводу: в течение всего фанерозоя
типы природной среды, существовавшие на континентах и шельфах, были во многом
сходны с существующими сейчас. Но пространственное положение их постоянно
менялось. Эти изменения вызывались местными поднятиями или прогибаниями поверхности, перемещениями плит земной коры
в более высокие или более низкие широты с различным климатом, а также
предположительно столкновениями континентов или излияниями магмы на
поверхность, в результате которых образовывались вулканические конусы или
обширные лавовые покровы. Моря,
побережья и возвышенности в ордовикское время. Для того чтобы понять, каким
образом изучаются природные условия прошлого, рассмотрим сильно отличающуюся
от современной обстановку, существовавшую на известной нам территории
Северной Америки в определенный момент времени в конце ордовика,
около 465 миллионов лет назад. Отложения, накапливавшиеся тогда на территории
современных штатов Нью-Йорк, Пенсильвания, Огайо и на юге Онтарио, хорошо
сохранились и во многих местах выходят на поверхность, оказываясь, таким
образом, легко доступными для изучения. При прослеживании их с запада на
восток устанавливается изменение их характера. В Огайо и западном Онтарио
породы этого возраста представлены довольно тонкими, почти горизонтально
залегающими слоями известняков с богатой ископаемой фауной беспозвоночных,
типичной для дна мелководного моря. Эти слои свидетельствуют о том, что эту
часть Северной Америки занимал мелководный морской бассейн, в котором обитало
большое количество разнообразных растений и животных. Общая картина,
вероятно, напоминала картину, изображенную на рисунке 37. Вода, очевидно, в
этом бассейне была чистой, так как известняки состоят главным образом из
кальцита; примесь глинистых частиц, которые могли бы загрязнять воду,
невелика.
Рис. 37. Природные условия юго-восточной части
современной области Великих озер в конце ордовика.
Пунктирными линиями на верхней поверхности блок-диаграммы показано положение
границ штатов и современные контуры Великих озер. Длина передней части блока
- около 1120 км Однако
по мере продвижения на восток мы обнаруживаем всевозрастающее содержание
глины в этих пластах, которое в конце концов достигает такой величины, что
породы переходят в аргиллиты. Далее появляются песчаные частицы, и аргиллиты
сменяются восточнее алевролитами, а затем песчаниками. В этом же направлении
увеличивается и мощность пластов. Эта смена типов пород сопровождается
изменением характера слоистости и исчезновением морских ископаемых
организмов. Согласно
нашей интерпретации, такие перемены объясняются тем, что при движении на
восток мы переходим из области древнего моря, пересекая побережье, в область
низменной пологой древней береговой равнины. Последняя была создана
процессами накопления аллювия, отложенного реками, которые текли с востока на
запад. Возрастание крупности зерен осадков в восточном направлении означает,
что верховья рек находились в пределах холмистой шш
низкогорной области к востоку от береговой равнины. Уклоны потоков должны
были быть настолько велики, чтобы сделать возможным перенос песчаных частиц. Сортировка
материала горных пород в пределах холмистой области осуществлялась химическим
выветриванием, как это было описано в пятой главе; не измененные
выветриванием частицы кварца выносились на запад, по направлению к морю. Все
эти частицы, соответствующие по крупности песку или алевриту, как и большая часть
глинистых частиц, отлагались в пределах береговой равнины. Далее по
направлению к морю вода, не содержавшая глинистых частиц, должна была быть
чистой и представлять благоприятную среду для обитания многих животных и
некоторых морских растений. Мы
почти не находим свидетельств существования песчаных пляжей в эпоху
формирования рассматриваемых отложений. Это не удивительно, так как часть
отложений, состоящих из песчаников, располагается дальше к востоку. Вблизи
побережья пески, которые могли бы образовать пляжи, почти полностью
отсутствовали, и берег был низменным и топким, напоминавшим современную
дельту реки Миссисипи. Действительно, берег к югу от Онтарио представляется
состоящим из целого ряда больших и малых дельт. Что касается растительности,
то нет никаких данных, указывающих на произрастание на суше в эти древние
времена каких-либо растений. Поэтому можно предполагать, что в ордовике береговая равнина представляла собой обширную
обнаженную илисто-песчаную поверхность, увлажненную и пересеченную мутными,
медленно текущими реками. Окажись там человек, он нашел бы эту обстановку
крайне непривлекательной. Холмы
и низкогорье, поставлявшие глинистые продукты выветривания коренных пород,
давно уничтожены. Сейчас на их месте располагаются выходы метаморфических
пород, простирание которых (с северо-востока на юго-запад), в общем, более
или менее параллельно береговой линии ордовикского моря. Холмы, являвшиеся
"предками" современных Аппалачских гор, занимали часть современной
территории Новой Англии. В то время, когда здесь происходило сжатие и
метаморфизм пород, зона земной коры, расположенная далее к западу, постепенно
прогибалась, образуя карман с большой мощностью отложений (как показано на
рис. 37). Такие прогибающиеся карманы мы встречаем на континентальных
шельфах. Географические
карты древних эпох. Реконструированная нами выше география древней эпохи
может не быть точной в деталях, однако она в достаточной степени основана на
фактах и позволяет правильно воссоздать облик древней поверхности Земли. Если
в нашем распоряжении имеется достаточное количество пластов осадочных пород,
то подобную реконструкцию древних природных условий можно сделать на любом
участке суши для одного или нескольких периодов времени, охарактеризованных
доступными для изучения осадочными породами. Совместными усилиями многих
современных геологов были созданы карты, показывающие географическую
обстановку Северной Америки и других континентов в различные прошлые эпохи, о
которых мы можем судить по геологическим разрезам. Точность этих карт
неодинакова, поскольку неодинаково количество информации, которое может быть
получено из геологических слоев. Для некоторых районов или промежутков
времени данные очень скудны или вообще отсутствуют - либо потому, что осадкона-копление в этих районах не происходило, либо
потому, что горизонты, необходимые для точных реконструкций, были
впоследствии уничтожены эрозией. На
картах показано постоянно менявшееся во времени распределение мелководных
морей, равнин и различных типов гор, а также вспышек вулканической активности
в том или ином районе. Рисунок каждой карты отражает постоянное
взаимодействие внутренних - эндогенных - и внешних - экзогенных - процессов.
Детали карт интересны главным образом для ученых, изучающих Землю. Для наших
целей достаточно сравнить две карты (рис. 38), показывающие строение
поверхности Северной Америки по данным реконструкций для середины девонского
и позднемелового времени. Эти геологические периоды разделены промежутком
времени более чем 200 миллионов лет.
Рис. 38. Слева: обобщенная карта Северной Америки в
среднедевонское время, показывающая положение морских проливов (штриховка) и
суши (белая). Для сравнения тонкими линиями показаны очертания современных
континентов. Справа: Северная Америка в позднемеловое время. Условные
обозначения те же Северная
Америка в палеозое. Если отвлечься от деталей и рассматривать лишь общую
картину, то карта среднедевонской эпохи (рис. 38, левый) может быть
использована для получения общего представления об условиях, существовавших
на территории Северной Америки в палеозое. Главной ее особенностью было
наличие двух узких и длинных внутренних морей, одно из которых располагалось
в районе современного восточного побережья, а другое гораздо дальше к западу.
Восточное море, показанное на этой карте, было одним из бассейнов,
существовавших на месте ордовикского морского бассейна, показанного на
рисунке 37. Хотя очертания его берегов менялись и по временам этот узкий
пролив полностью высыхал, море снова и снова занимало свои прежние границы,
что, по крайней мере отчасти, объяснялось слабым, но постоянно
возобновлявшимся прогибанием земной коры в этой
области. Временами
этот прогиб, по крайней мере частично, подвергался боковому сжатию, так что
выполнявшие его осадки сминались в складки (рис. 37). Возможно, это явление
вызывалось сближением одного континента с другим, занимавшим тогда положение
вблизи нынешнего восточного побережья. Но до самого конца палеозойской эры,
на протяжении более чем 300 миллионов лет, море продолжало вновь и вновь
возвращаться на то же самое место, пока в результате заключительного сжатия
осадки, выполнявшие прогиб, не были смяты, образовав Аппалачские горы, и весь
район стал сушей. В настоящее время, после того как эти горы в течение 250
миллионов лет (всего мезозоя и кайнозоя) подвергались эрозии, удалившей
огромные массы выветрелых пород, они все еще
существуют как остатки некогда величественных сооружений. Наклонно залегающие
слои песчаников и алевролитов устойчивы против эрозии и образуют длинные
гребни, в то время как понижения между гребнями выработаны в менее устойчивых
известняках и аргиллитах (фото 21).
Фото 21. Аппалачские горы южнее Гаррисберга,
Пенсильвания. Хребты на заднем плане вытянуты в прямую линию и почти
параллельны друг другу, гребень на переднем плане образован резко изогнутым
пластом пород. Прорезавшая его по диагонали эрозия создала зигзагообразные
формы Возвращаясь
к морям, существовавшим до образования Аппалачских гор, отметим, что жизнь в
море развивалась в течение более 300 миллионов лет палеозойской эры и
появилось множество рыб. Окружающая суша, в ордовике
еще обнаженная, покрылась примитивной растительностью, появились наземные
животные, образовавшие большое количество видов. Подобной
же была история и другого узкого морского бассейна Северной Америки. Он
занимал зону, неоднократно подвергавшуюся прогибанию,
где так же отступало и наступало море и в которой накапливались мощные толщи
осадков. В мезозое море просуществовало недолго, исчезнув уже в триасовом
периоде. Северная
Америка в меловое время. Внимательно рассмотрев карту среднедевонского
времени и приобретя, таким образом, некоторый опыт, мы можем обратиться к
карте позднемеловой эпохи (рис. 38, правый). На этой карте мы видим Северную
Америку, какой она была 70-75 миллионов лет назад. Море покинуло район
Аппалачей перед концом, палеозоя. Атлантический берег находился восточнее
современного, за исключением полуострова Флорида, большая часть которого была
затоплена. Огромные пространства восточной части континента представляли
собой сушу. На карте резко выделяется довольно широкий пролив, соединявший
водное пространство, находившееся на месте современного Мексиканского залива,
с Северным Ледовитым океаном. Этот пролив разделял Северную Америку на две
части. В районе Тихоокеанского побережья море покрывало большую часть
современной прибрежной суши. Существовавшая тогда суша в основном была
низменной и незначительно превышала уровень моря; обширные прибрежные
пространства изобиловали болотами. Климат был мягче современного. Суша была
сплошь покрыта растительностью, которая носила экзотический по сравнению с
современной растительностью характер. Среди наземных животных преобладали
рептилии, главным образом динозавры, процветавшие в условиях мягкого климата
низменностей. В морях обитало огромное количество рыб и разнообразных
беспозвоночных, а воздушным пространством завладели летающие рептилии и
некоторые виды птиц. Реконструкция
природных условий Мы
рассмотрели реконструированные природные условия, существовавшие на востоке
территории Северной Америки в ордовике. Мы
ознакомились также с картами поверхности Северной Америки, какой она была в
два различных отрезка времени в прошлом - в девонскую и меловую эпохи. На
каких же данных, полученных при изучении осадочных пластов, основаны эти
реконструкции? На указанных картах представлена история, и составившие их
геологи в такой же мере являются историками, как и те, кто восстанавливает
деяния людей в прошлом. Для
историка большой удачей является находка письменных документов,
свидетельствующих о событиях, отстоящих от нас более чем на несколько тысяч
лет. Хотя ученый, занимающийся историей Земли, совсем не располагает
письменными документами, он может найти множество естественных документов в
геологических пластах, часто содержащих ископаемые остатки. Он может
пользоваться также методом радиометрического датирования, с помощью которого
он, как мы видели, составил надежную хронологическую шкалу. Примеры
интерпретации природных условий. Когда ископаемые моллюски, показанные на
фото 22, были собраны, определены и сопоставлены с родственными, ныне
живущими видами, оказалось, что они представляют собой организмы, обитавшие
не в открытом море, а в защищенных солоноватых водах, там, где реки впадают в
море. Тонкие, почти параллельно залегающие слои ила, в которых они найдены,
также могут указывать на условия устьевой части реки - эстуария. Путем
корреляции установлена принадлежность этих ископаемых остатков к началу
плейстоцена, возможно, ко времени около двух миллионов лет назад. Этот
довольно молодой возраст подтверждается состоянием осадков - ил еще не
превращен в твердый алевролит - и самых раковин, еще не подвергшихся
окаменению. Хотя место находки ископаемых сейчас расположено в 16 километрах
от моря и почти на 60-метров выше его уровня, совершенно ясно, что в то время,
когда жили эти моллюски, здесь находилась приливная зона, возможно, в
пределах затопленной морем долины реки Тибр. С тех пор эта часть западного
побережья Италии была приподнята, что вызвало отступание моря и обсыхание ранее частично затопленного прибрежного района.
Напротив, на фото 19 изображен участок в пределах штата Вайоминг,
расположенный вдали от моря в средней части континента. На этом участке была
обнаружена передняя конечность животного, настолько огромная, что она должна
была принадлежать динозавру. Он относился к роду Diplodocus
и был подобен животному, изображенному на фото 45. Будучи рептилией, этот
динозавр должен был жить в сравнительно мягком климате. Строение его скелета
показывает, что он ходил на четырех ногах и питался растениями, очевидно
мягкими и в какой-то степени сходными с водорослями. Породы, в которых были
найдены некоторые остатки, относятся к позднеюрскому времени (возраст их
предположительно 140 миллионов лет) и представлены в основном тонкозернистым
аллювием и отложениями, характерными для болот и замкнутых водоемов.
Фото 22. Слоистый ил, содержащий ископаемые моллюски
различных видов. Справа от середины снимка помещен карандаш длиной 12,5
сантиметров. Бассейн реки Тибр ниже Рима По
этим данным был сделан вывод, что район Вайоминга в это время представлял
собой ряд обширных речных равнин с многочисленными сообщающимися болотами и
озерами, с богатой растительностью и теплым климатом - сочетание условий,
напоминающее бассейн современной реки Амазонки. Высота местности над морем
была очень невелика; это можно легко установить из того, что в позднеюрское
время внутреннее море, подобное изображенному на рисунке 38, правом, но менее
обширное, затопляло западную часть района Вайоминг несколько западнее
участка, изображенного на фото 19. Если
слои серии, в которых были заключены кости упомянутого динозавра, отлагались
на низменной равнине вблизи уровня моря, в мягком и влажном климате, то с тех
пор условия среды сильно изменились. Теперь место находки этих ископаемых
остатков расположено почти на 2,5 километра выше уровня моря, в семиаридном климате с разреженной растительностью и очень
холодной зимой. Здесь проявились эндогенные процессы, в результате которых
обширные площади на западе Северной Америки испытали сводовые
поднятия, начавшиеся в конце мелового периода и продолжавшиеся в течение
большей части кайнозойской эры. Таким
образом, из частичных и обрывочных данных, полученных главным образом при
изучении осадочных отложений, постепенно складывается картина медленно
изменяющейся в течение последних 600 миллионов лет природы Земли. Частично
эти сведения дает анализ физических характеристик пластов, но более важную
роль играет изучение ископаемых остатков. Из года в год мы получаем все
больше сведений, позволяющих нам представить более ясную картину, но пройдет
еще много времени, прежде чем мы сможем прекратить заполнять пробелы в
исторических данных домыслами. |