ЭКОЛОГО-ГИДРОГЕОЛОГИЧЕСКОЕ ПРЕДПРИЯТИЕ
"ЭКОМОНИТОРИНГ"
СРЕДНЕУРАЛЬСКАЯ ГИДРОГЕОЛОГИЧЕСКАЯ ЭКСПЕДИЦИЯ
|
620014
г.Екатеринбург ул.Вайнера, 55 (Уралнедра), каб. 513 |
тел. 257-20-06, 219-39-08 факс 257-20-06 |
|
|
|
Глава одиннадцатая. Геологическая история растений Наш
обзор развития биосферы в фанерозое логично начать
с растений. Растения не только прямо или опосредствованно служат пищей
животным; летопись ископаемых остатков показывает, что растения раньше
животных покинули море, эту колыбель всех форм жизни. Следовательно, мы
сможем легче проследить историю развития животных, если сначала опишем
растительный покров континентов и развитие растительности до наших дней.
Летопись ископаемых растений, особенно в своей ранней части, изобилует
довольно большими пробелами. Это не удивительно, потому что условия для
сохранности ископаемых остатков на суше хуже, чем на дне моря, а большинство
ископаемых растений росло именно на суше. Такие пробелы затрудняют
установление эволюционных связей между группами растений. Несмотря на это,
если не вдаваться в детали, можно наметить общую историю, основанную на
известных к настоящему времени растительных остатках. Структура
и физиология растений Сосудистая
система. Краткий обзор особенностей современных растений - основных черт их
строения и физиологии - показывает результат развития растительного мира.
Большинство растений, особенно высших, отличается характерными чертами
строения; все они, в частности, имеют: 1) длинный, обычно вертикальный и
ветвящийся стебель, в большинстве случаев жесткий; 2) корни, хорошо
закрепленные в почве; 3) в верхней части - зеленую листву. Самый важный
процесс - фотосинтез - происходит в листьях, безразлично, широкие это листья,
иглы или еще какие-нибудь. Клетки листьев синтезируют хлорофилл
("растительную зелень"), зеленый пигмент, который поглощает
солнечный свет и, используя его энергию, может создавать питательные вещества
из углекислого газа атмосферы и воды, которую растение извлекает из почвы.
Через поры листьев поступает углекислый газ, происходит реакция фотосинтеза
(химическое восстановление кислорода) и в результате образуются сахар, вода и
свободный кислород. Вода,
которая нужна для фотосинтеза, поступает снизу, из почвы, по системе корней,
через стебель и ветви. Теперь мы понимаем, что корни, стебель и ветви созданы
не просто для того, чтобы нести на себе листья. Это также и водоподводящая
система каналов, по которой вода в достаточных количествах подается даже на
вершину самого высокого дерева. Очень высокие хвойные деревья на
тихоокеанском побережье Северной Америки могут подтягивать воду на высоту до
120 метров над землей. В
действительности вода, которая подтягивается вверх, к листьям, это не просто
вода; это сок деревьев, водный раствор. В одеревеневшей ткани живого дерева
находятся вертикальные цепочки мертвых клеток. У некоторых видов деревьев
верхние и нижние стенки этих клеток растворены - как если бы в высоком здании
пришлось уничтожить перекрытия в целом ряде комнат, расположенных одна над
другой, чтобы соорудить шахту для лифта. Цепочки этих мертвых клеток
заполнены соком. Они образуют трубочки, по которым жидкость, поступающая из
земли, подтягивается от корней к живым листьям. Из листьев вода постоянно
испаряется в атмосферу через поры. Эта потеря влаги снижает давление внутри
листьев и тонких веток и заставляет сок, заполняющий систему трубок,
подтягиваться через стенки клеток к листьям, чтобы возместить недостаток жидкости.
Таким образом всасывающее усилие передается по всей системе трубок, включая
корни, которые в свою очередь начинают более активно поглощать влагу из
почвы. Вся эта система "сосудов" представляет собой своего рода
насос, действующий весьма эффективно. Эта
система, рассматриваемая в более широком плане, представляет собой часть
круговорота воды (рис. 4). В таком круговороте часть осадков просачивается в
землю, поглощается корнями растений, поднимается к листьям и с их поверхности
испаряется в атмосферу, из которой выпадает затем в виде дождя. Но часть воды
остается в растениях. В процессе фотосинтеза она соединяется с углекислым
газом, который проникает в растение из атмосферы через поры листьев, и
образует питательные вещества и ткани растения. Для
образования тканей растения требуется большое количество воды и углекислого
газа. Например, на 0,4 га кукурузного поля произрастает около 10 000
растений, которые дают около 3,5 м3
зерна (семян кукурузы). Для образования этого количества семян (мы не считаем
стебли, корни, листья и стержни початков) растениям требуется около 9 т
углекислого газа, из которого они выделяют и используют 2,5 т углерода, а
освободившийся кислород возвращают в атмосферу. Для этого растения должны
переработать 21 000 т воздуха, то есть более 2 т на каждое растение. В то же
время растения должны извлечь из почвы и перекачать в листья 1 892 500 литров
воды, что составляет около 190 литров на каждое растение. Этот
процесс происходит на каждом участке кукурузного поля ежегодно. То же самое
происходит на площадях, засеянных пшеницей и другими зерновыми. Такие же
процессы, хотя и происходящие более медленно, наблюдаются на площадях,
занятых деревьями, дающими фрукты или орехи или использующимися как топливо и
строительный материал. Представив себе огромные пространства лесов, садов,
пастбищ и обрабатываемых полей, безразлично, используемых человеком или нет,
мы до некоторой степени можем понять, какую огромную роль растения, входящие
в состав биосферы, играют в круговороте воды, а также в процессах
выветривания (физического и химического), образующих часть круговорота горных
пород. Система
размножения. Изучением способа размножения живых организмов, животных или
растений, никогда не следует пренебрегать, потому что успешное
воспроизводство организма - это один из важнейших факторов его успешной
эволюции, залог способности выживать в меняющихся (к лучшему или худшему)
условиях. Поэтому, рассмотрев способы размножения современных растений, мы
можем узнать что-то новое и о геологической истории. Небольшая
часть ныне существующих растений размножается вегетативно. При вегетативном
размножении у растения развиваются отростки, луковицы или побеги, которые
укореняются и становятся самостоятельными растениями, точно воспроизводящими
своего родителя. (Мы употребляем это слово в единственном числе, так как в
этом случае родитель только один.) При таком размножении сочетания различных
генов не происходит; потомок не приобретает никаких новых свойств, которые
могли бы послужить ему для приспособления к новым условиям среды. Половое
размножение, напротив, допускает сочетание различных свойств, унаследованных
от различных особей, и если условия среды благоприятствуют развитию
возникающих признаков, то эволюция ускоряется. В своей основе цикл полового
размножения бессеменных сосудистых растений
включает две фазы. В первую фазу возникает маленькое безлиственное растение,
продуцирующее сперматозоиды, или яйцеклетки, или то и другое одновременно.
Сперматозоиды от мужского растения перемещаются в воде (присутствующей хотя
бы в виде тонкой пленки) и оплодотворяют яйцо женского растения, находящегося
поблизости. Из оплодотворенного яйца развивается новое растение,
превосходящее по размерам растения первой фазы и имеющее листья. Во вторую
фазу это большее растение образует споры - клетки, служащие для размножения и
не требующие оплодотворения. Они уносятся и рассеиваются ветром, укореняются,
и из них развиваются мелкие безлиственные растения первой фазы. Таким образом
завершается цикл. Как мы видим, один момент данного цикла требует присутствия
воды; на ранних этапах истории наземных растений это ограничивало их
распространение, так как размножение могло происходить лишь в определенных
природных условиях. Появление
семян устранило это затруднение и таким образом усовершенствовало процесс
размножения. При наличии семян отпала необходимость движения сперматозоидов к
яйцеклетке вводе. Семя - это не просто изолированная клетка. Оно включает
множество клеток, образующих в совокупности корень, стебель и один-два листка, "упакованных" в питательное
вещество и снабженных оболочкой, защищающей их от механического и химического
воздействия. Кроме того, когда семя начинает свое существование, оно уже
содержит оплодотворенную яйцеклетку. Оплодотворение происходит тогда, когда
яйцеклетка еще не отделилась от родительского растения, которое и питает семя
на начальных стадиях роста. К тому времени, когда семя отделяется, оно уже
представляет собой миниатюрный организм, снабженный
"приспособительными" устройствами. Этот организм может сохраняться
живым до тех пор, пока не укоренится и не начнет свою жизнь как
самостоятельное растение. Благодаря указанным свойствам он имеет гораздо
большие шансы выжить по сравнению с бессеменными
растениями, что в свою очередь означает большее разнообразие возможных сред
обитания и лучшую конкурентную способность данных видов. Значение таких
преимуществ показывает вся история развития растений. Древнейшие
морские растения Эволюция
растений начинается с морских водорослей (см. табл. 2). Самые древние из
известных на Земле ископаемых остатков (возраст которых около трех миллиардов
лет) представляют собой наиболее примитивные растения - сине-зеленые
водоросли. Почти два миллиарда лет такие растения обитали в морях вместе с
другими одноклеточными организмами. Но, как свидетельствуют находки
ископаемых в Австралии, около одного миллиарда лет назад существовали и
зеленые водоросли. Наличие зеленых водорослей показывает, что к этому времени
растения приобрели два важнейших свойства в отличие от их предков - более сложное
строение и способность к половому размножению. Последнее свойство мы уже
рассматривали. Поскольку каждый организм-потомок происходил уже не от одного,
а от двух родителей, он обладал новой комбинацией генетического материала,
что обусловило большую эффективность эволюции. Усложнение
структуры хотя и не было столь заметным усовершенствованием, также имело
существенное значение для будущего. Когда первичные одноклеточные растения
приобрели вторую клетку, а затем еще большее число клеток, их форма изменилась
от почти шарообразной до линейной или неправильной. Возможное объяснение
этого факта заключается в том, что любая несферическая форма имеет большее
отношение площади к объему по сравнению с шаром. Таким образом, приобретение
растением линейной или неправильной формы означало увеличение площади
соприкосновения с морской водой, то есть площади поверхности, поглощавшей
углекислый газ и солнечные лучи, необходимые для питания растений. Когда
создались неправильности формы, из них могли развиться органы, выполняющие
различные функции. И это должно было случиться раньше, чем водоросли могли из
воды выйти на сушу. Заселение
суши растениями До
сих пор известны только две находки очень древних ископаемых наземных
растений. В верхнесилурийских слоях Центральной Европы были найдены остатки
безлистных растений длиной до 10 сантиметров; это самые древние из известных
нам наземных растений. Вторая находка была сделана в Австралии, ц слоях раннедевонского возраста. Она представляет собой
окаменевшие стебли (до 25 сантиметров длиной) растения, очень близкого к
современным плаунам Lycopodium (фото 23), стебель
которых стелется по земле и образует вертикальные отростки. Lycopodium - это не только наземное растение, это также и
сосудистое растение. Наличие ископаемых растений такого типа означает не
только существенный прогресс эволюции, но также и крупный географический
сдвиг - выход растительной жизни из моря на сушу.
Фото 23. А. Древнейшее из известных нам наземных
растений. Ископаемый отпечаток растения рода Baragwanathia
в нижнедевонских слоях юго-восточной Австралии. Совершенно
ясно, что к позднесилурийскому времени растения уже заселили сушу. Но сколько
времени занял этот процесс? Между появлением древнейших известных нам морских
зеленых водорослей (около одного миллиарда лет назад) и древнейших наземных
растений, обнаруженных в позднем силуре Европы (приблизительно 430 миллионов
лет назад), прошло около 570 миллионов лет. Это действительно большой отрезок
времени, почти равный продолжительности всего фанерозоя.
Хотя мы и признаем, что заселение суши мягкими морскими водорослями и
развитие у них сосудистой системы были их замечательным!
"завоеваниями", мы не должны забывать, что эти растения имели в
своем распоряжении очень много времени, достаточно много для миллионов
"проб и ошибок". Если вспомнить, что за такое длительное время даже
ничтожная на первый взгляд вероятность почти наверное реализуется, то мы
вправе усомниться в том, что водорослям действительно потребовалось так много
времени (570 миллионов лет) для завоевания суши. Может быть, где-нибудь
существуют кембрийские или даже более древние слои, содержащие остатки
наземных растений, и если это так, то когда-нибудь они, вероятно, будут
найдены и определены. И тогда сократится большой пробел в известной нам
истории развития растений.
Фото 23. Б. Близкородственное ему современное
растение, плаун Lycopodium lucidulum
с побегами длиной 10-12 см Путь
из моря на сушу. Пытаясь заполнить этот пробел логическими рассуждениями, мы
можем представить, каким образом произошел этот великий переход растений из
моря на сушу. Водоросли, жившие в море, выделяли свободный кислород, который
поступал в атмосферу, приближая постепенно ее состав к современному. Кислород
(См. примечания к главе седьмой, стр. 106) создал экран, защищающий организмы
от смертоносного излучения Солнца, и таким образом позволил им переселиться
из моря на сушу, не подвергаясь опасности немедленной гибели. Этими
переселенцами были растения, очевидно, зеленые водоросли. Они сами
подготовили условия для этого переселения, выделяя в ходе многих лет процесса
фотосинтеза огромные массы свободного кислорода, который превратил атмосферу
Земли в защитный экран. Естественно
предположить, что зеленые водоросли расселялись вдоль побережий и проникали в
устья рек. Через миллионы лет они, вероятно, распространились вверх по рекам,
перейдя из соленой морской воды через солоноватую воду устьев рек в пресную
речную. Мы можем представить себе, что временами медленные поднятия на
некоторых участках суши преграждали путь рекам и создавали озера. С течением
времени озера могли неоднократно изменять уровень и даже высыхать, а потом
снова возникать. Непрекращающееся взаимодействие экзогенных и эндогенных
процессов могло принимать тысячи различных форм. Конечно,
мы не знаем, сколько видов водорослей прошло этот путь. Но по крайней мере
один из них выжил в условиях пересыхающего водоема. Живя в воде, водоросли
поглощали пищу всей своей поверхностью. Но когда они стали
"выходить" на сушу, им пришлось отказаться от этого простейшего
способа питания. Первый шаг в этом направлении мог быть вызван опытом жизни в
условиях временного пересыхания, возможно, в устьевой части речной долины во
время отлива. У растений развился подобный коже покров, защищавший их от
высыхания; в то же время этот новый покров делал возможным поступление воды в
растения и ее диффузию. Одновременно
неровности тела растения должны были преобразоваться в примитивные органы со
специализированными функциями. Один такой выступ, возможно нитевидный, мог
использоваться для прикрепления к грунту, а также для поглощения воды и
растворенных питательных веществ из грунта. Должны были также развиваться
дополнительные сосудистые ткани, служившие опорой для тянущегося вверх
растения. Иначе говоря, постепенно растение стало уже существенно отличаться
от прежней мягкой водоросли. Оно становилось примитивным наземным растением,
обладающим подобием корня, стеблем и простейшей сосудистой системой. Однако
голые зеленые стебли и тонкие ветви, лишенные листьев, все еще напоминали
водоросли. К
позднесилурийскому времени (возможно и раньше) такая стадия водорослеобразных растений была достигнута и пройдена.
Как только закончился сложный процесс выхода растений на сушу, его
естественным результатом явилось расселение и приспособление их к различным
местным условиям, что должно было протекать сравнительно легко. Основные
предпосылки для этого - наличие почвы, воды, минеральных питательных веществ
в почве и углекислого газа в воздухе - уже существовали. Поверхность Земли,
напоминавшая поверхность Луны, начала одеваться зеленым покровом. Древняя
наземная растительность; бессеменные растения.
Некоторое представление о древнейшей растительности дают остатки окаменевших
растений, встречающиеся местами в пластах горных пород, например в речных
отложениях первой половины девонского времени. Эти растения принадлежат к
нескольким различным родам, но все они относятся к примитивным группам,
впоследствии исчезнувшим. Большая часть их имела стелющиеся по земле стебли с
отростками, поднимавшимися на несколько десятков сантиметров, без листьев или
с зачатками листьев. Такие растения не имели семян. По найденным ископаемым
остаткам были сделаны реконструкции, одну из которых вы видите на фото 24.
Фото 24. Реконструкция раннедевонского ландшафта, на
котором показаны растения нескольких родов, найденные в ископаемом состоянии
в различных районах земного шара. Для масштаба показана дорожная сумка По
мере того как растения приспосабливались к различным условиям, они постепенно
дифференцировались, образуя все большее количество видов. К концу девона, то
есть 360 миллионов лет назад, растительность стала иметь уже совсем другой
облик. Выделялись папоротниковидные растения, предки настоящих папоротников.
Они не давали семян; некоторые из них достигали 12 метров в высоту и около
одного метра в диаметре ствола у основания. Представив себе эти размеры, мы
можем сказать, что первые леса на Земле начали возникать в позднедевонское
время. Кроме древовидных растений, в этот период времени существовали крупные
хвощи и плауны, потомки более мелких видов, живших ранее в девоне. На фото 25
изображена другая реконструированная картина, предположительно показывающая
облик позднедевонского ландшафта.
Фото 25. Реконструкция ландшафта позднедевонского
времени, на котором показаны настоящие папоротники (П),
примитивные лепидодендроны - "чешуйчатые деревья" (Ч) и меньшие по
размеру растения Позднепалеозойская
флора; семенные растения. Следующим важным событием в истории растений было
появление семян, на что ясно указывают ископаемые из каменноугольных пластов.
Семена эти были довольно примитивными: они не имели оболочки и развивались на
плодолистиках. Яйцеклетка семени оплодотворялась сперматозоидами того же
самого растения. Среди древнейших растений, размножавшихся семенами, были
семенные папоротники. Современные потомки других древнейших растений,
размножавшихся семенами, - хвойные (сосна, ель, тсуга) - до сих пор
размножаются таким же образом. Как и семенные папоротники, хвойные и другие
подобные растения принадлежат к группе так называемых голосеменных, и все они
имеют семена самого примитивного типа. Однажды
появившись, голосеменные растения уже не обязательно должны были держаться
вблизи увлажненных мест, потому что для их способа размножения не требовалось
даже тонкой пленки воды. Они занимали возвышенности, горы и сухие участки,
ранее лишенные растений, и в результате этого зеленый растительный покров
распространился по Земле еще больше. Очевидно,
раз голосеменные существовали в течение 350 миллионов лет и до сих пор еще
очень многочисленны, их система размножения действует нормально. Но для того,
чтобы она действовала, каждое голосеменное растение всегда должно производить
большое количество семян и еще больше - пыльцы (пылевидных мужских половых
клеток) для опыления. Размножение голосеменных зависит от ветра, разносящего
их семена, но при этом многие семена попадают в неподходящие условия, - в
воду, в пустыни, в места, слишком холодные для прорастания. Голосеменные
растения затрачивают много энергии на производство семян и пыльцы, большая
часть которых погибает. Впоследствии растения преодолели эту трудность, найдя
более экономичный метод, но это произошло уже почти на 200 миллионов лет
позднее, в меловой период. Представьте
себе условия, которые существовали в течение данного большого промежутка
времени. На континентах в позднекаменноугольное
время преобладали обширные низменности с мягким климатом. В обширных болотах
обильно произрастала пышная растительность; болотная вода покрывала отмершие
части растений, предохраняла их от разложения и способствовала их сохранности
вплоть до спрессования и превращения в уголь. Таким
образом, в нашем представлении каменноугольная растительность - это главным
образом богатая растительность влажных низменностей. Об обстановке,
существовавшей в это время на холмистых возвышенностях, мы знаем мало, потому
что более крутые склоны возвышенностей и отсутствие, как правило, на них
стоячих вод не способствовали сохранности отмерших растений. Болотистые
низменности могли выглядеть так, как это изображено на фото 26, показывающем влажный
лес. В таком лесу водилось много животных, подобных саламандрам, и насекомых.
Такой тип болот, конечно, встречался не только в позднекаменноугольное
время, он существовал и во все последующие эпохи. В настоящее время он часто
встречается по берегам Мексиканского залива и вдоль Атлантического побережья
Соединенных Штатов. Таковы в общих - чертах современные болота, хотя,
конечно, большая часть растений болот принадлежит теперь к совершенно другим
видам, чем в каменноугольный период.
Фото 26. Реконструкция болота позднекаменноугольного
времени с обычной для таких болот пышной растительностью. П
- папоротники, X - хвощи, Ч - лепидодендроны -
"чешуйчатые деревья", СП - семенные папоротники; видна также
большая стрекоза. Некоторые хвощи достигали в высоту 30 м Мезозойская
растительность. Следующая из наших реконструкций относится к триасовому
периоду и характеризует район национального парка "Каменный лес" в
восточной Аризоне В действительности это совсем не лес. Это остатки леса,
росшего Гдругом месте; они представляют собой
скопление стволов деревьев, заключенных в толщу аргиллитов, из которой они
сейчас постепенно высвобождаются в результате процессов выветривания (фото
27). За время долгого пребывания в толще породы стволы окаменели, их
клеточная структура была замещена кремнеземом и превращена в агат -
разновидность кварца. Стволы принадлежали хвойным деревьям (конечно,
голосеменным), родственным современным соснам. При жизни деревья достигали
30-60 метров высоты, а стволы их имели толщину около двух метров. Мы знаем,
что они росли не там, где находятся сейчас, потому что их поверхность носит
следы истирания, а все ветви и корни обломаны. Это - плавник, принесенный в
половодье откуда-то с возвышенностей в верховьях реки и погребенный в тонких
аллювиальных отложениях. Он свидетельствует о том, что уже 220 миллионов лет
назад на западе Северной Америки росли величественные рощи хвойных деревьев, точно
такие же, как и те, которые значительно позже увидели европейцы, впервые
проникшие на континент.
Фото 27. Огромные стволы триасовых хвойных деревьев,
окаменевшие и превратившиеся в кварц. Национальный парк "Каменный
лес", Аризона Фото
28 представляет собой реконструкцию растений, характерных для мезозойской
эры. Среди них преобладают две группы голосеменных - хвойные и цикадовые, а
также родственные им виды. Хвойные включали виды, подобные соснам и
кипарисам, а также гинкго. Цикадовые, вероятно,
развились из семенных папоротников. Их внешний облик напоминал пальмы, хотя
эволюционной связи между этими группами нет; изящная крона из папоротниковидных
листьев увенчивала ствол короткий, шарообразный или бочонковидный,
а иногда высокий. Некоторые цикадовые сохранились до наших дней. Бессеменные растения, в первую очередь папоротники и
хвощи, в мезозойское время играли менее заметную роль по сравнению с этими
голосеменными.
Фото 28. Реконструкция общего вида растительности,
типичной для мезозойской эры (только голосеменные и бессеменные
растения). 7. Растения, родственные цикадовым. 2 Растения, родственные
хвощам. 3. Папоротники. 4. Хвойные. Другие мезозойские ландшафты показаны
также на фото 40, 43, 44, 45, 47 Покрытосеменные
растения Цветы
и плоды. Длительная история растений ознаменовалась еще одним эволюционным
усовершенствованием - появлением цветковых растений, или покрытосеменных. Это
название дано им потому, что в отличие от голосеменных, у которых ничем не
защищенные семена росли прямо на листьях; у этих растений семена, поодиночке
или несколько вместе, заключены в особую оболочку, образующую внешнюю часть
плода (например, яблоко). Кроме того, покрытосеменные имеют не только плоды,
но и цветки. И то и другое приспособление обеспечивают лучшее размножение.
Яркие цветки привлекают опыляющих их животных, например пчел, которые
переносят пыльцу - мужские половые клетки; у более примитивных растений
перенос пыльцы зависел от ветра. Плоды также привлекают животных, которые
поедают их и таким образом разносят содержащиеся в них семена. Цветы,
плоды и другие усовершенствования позволили покрытосеменным растениями
заселить те участки суши, которые еще оставались безжизненными, - участки с
наиболее суровыми условиями существования. Этот процесс можно сравнить с
покорением человеком вершины Эвереста, которое стало возможным лишь после
того, как был создан ряд специальных приспособлений, в том числе кислородные
приборы. Сейчас покрытосеменные составляют более 95% существующих видов
сосудистых растений. Мы
не знаем, когда впервые появились покрытосеменные. Судя по ископаемым
остаткам, они определенно встречаются в меловых слоях и, возможно, в юрских и
даже в триасовых (остатки, содержащиеся в последних слоях, напоминают
покрытосеменные растения, но у них не сохранились части, которые точно
указывали бы на это). Во всяком случае, к середине мелового периода, около
100 миллионов лет назад, 90% видов растений, определенных в ископаемом
состоянии, относились к покрытосеменным. К этой же группе относится большая
часть современных лиственных деревьев, плодовых кустарников, овощей и цветов,
трав и злаков. Список этих растений довольно внушителен, так как появление
покрытосеменных знаменует переход от "средневекового" к
современному растительному миру. Уже позднемеловые флоры имели современный
облик, а существовавшие в начале кайнозоя (фото 48) деревья и другие растения
показались бы современному человеку, если бы он их увидел, знакомыми. Покрытосеменные
растения и климат. Наиболее заметное различие между покрытосеменными и более
ранней растительностью заключается в том, что покрытосеменные деревья и
кустарники повсюду, за исключением тропической зоны, являются листопадными.
После созревания семян они теряют листья, фотосинтез прекращается, и они на
время, иногда на несколько месяцев, "засыпают". На основании этого
ученые предположили, что появление покрытосеменных, по крайней мере отчасти,
явилось результатом изменения климата континентов, повлиявшего на процесс
эволюции. Если в мезозое климат приобретал все более резко выраженный
сезонный характер, причем сезоны становились все более холодными и сухими, то
опадение листьев и прекращение жизненной активности было бы вполне
целесообразным. Обдумывая это предположение, мы должны вернуться к рисунку 27
и посмотреть, как происходило перемещение плит земной коры и движущихся с
ними континентов в течение всего мезозоя. Северная Америка и по крайней мере
европейская часть Евразии двигались к северу в более высокие и более холодные
широты. Вполне возможно, что это предположение может объяснить появление
листопадных видов растений в юрский и меловой периоды. Это не более чем
предположение. Наука движется вперед, пытаясь связать воедино самые
разнообразные данные о прошлом Земли. Именно так были установлены связи между
отдельными событиями долгой истории Земли.
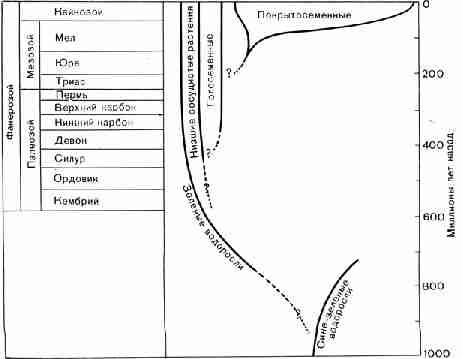
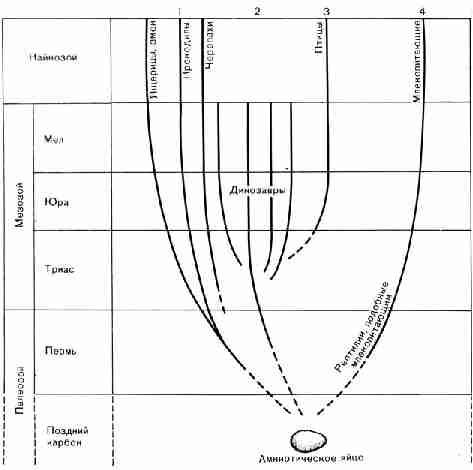
Рис. 39. Основные этапы геологической истории
растений Роль
трав и злаков. Если судить по времени установления господства
покрытосеменных, то растительность Земли приобрела современный облик еще до
конца мезозойской эры, однако история развития растений на этом не кончается.
Внутри самой группы покрытосеменных происходили существенные перемены,
начавшиеся перед концом мелового периода и продолжавшиеся в течение большей
части кайнозоя. Наибольший интерес для нас представляет развитие злаков. Многие
обитатели современных городов представляют себе травы только в виде газонов
или площадок для гольфа. Но травы, в частности злаковые, - это
самостоятельная группа покрытосеменных, которая приспособлена к жизни в
засушливом или сезонно-засушливом климате. Родина злаковых трав - сухие
равнины и степи с редким древостоем, называемые саваннами. Карта
растительности (рис. 35) показывает, что почти вся западная часть центра
Северной Америки представляет собой природную злаковую степь, не говоря уже
об огромных площадях разреженной травянистой растительности в пустынных и
полупустынных областях дальше к западу от штата Вашингтон до Мексики. Возможно,
благодаря распространению сухих климатов злаковые травы занимали все большую
и большую площадь в течение кайнозоя. Имея съедобные стебли, листья и семена,
злаковые травы являются идеальной высококалорийной пищей для животных,
которые приспособлены к их поеданию. И как мы вправе были ожидать, быстро
развивавшиеся в раннем кайнозое млекопитающие действительно поедали траву в
большом количестве. Путем естественного отбора они все лучше и лучше
приспосабливались к этому процессу. Форма их зубов и даже конечностей во
многом определялась травяным рационом. Хлебные
злаки - это особый вид злаковых трав, в котором концентрация питательных
веществ больше, чем в обычных злаках. Люди открыли высокую пищевую ценность
хлебных злаков десять тысяч лет назад или еще раньше. Вскоре после этого люди
начали сводить и расчищать леса, чтобы иметь возможность возделывать хлебные
злаки. Еще позднее они начали повышать урожайность злаков путем отбора семян
и искусственного выведения растений, а также путем применения удобрений и
орошения. Сейчас большинство людей питается главным образом злаками: рисом,
пшеницей, кукурузой и другими зерновыми. Десять тысяч лет - это короткий
промежуток времени в истории Земли, чтобы питание повлияло на строение
человеческого тела. Прежде
чем мы перейдем к рассмотрению эволюции животного мира, целесообразно
восстановить основные события истории развития растений, помня о том, что без
растений животный мир не смог бы успешно развиваться на суше. В результате
изучения ископаемых остатков можно установить следующие основные моменты в
развитии растений: 1.
Выход на сушу морских водорослей; примитивные наземные растения. 2.
Бессеменные растения. Папоротники, хвощи. 3.
Появление семян. Голосеменные. 4.
Появление покрытосеменных, характеризующихся наличием цветов и плодов. 5.
Развитие злаковых трав и позднее хлебных злаков. Эволюционные
соотношения между основными группами растений и хронологическая шкала истории
развития растений приведены на рисунке 39. Глава двенадцатая. Беспозвоночные и рыбы Основы
истории развития животных Геологическая
история развития растений дает хорошую основу для изучения истории животных
благодаря тесным биохимическим связям этих двух групп организмов: растения обеспечивают
животных пищей и свободным кислородом, а животные поставляют необходимый
растениям углекислый га Это взаимоотношение позволяет считать, что обе группы
oрганизмов развивались более или менее параллельно.
В предыдущей главе мы прочли, что эволюция некоторых животных происходила под
влиянием определенных видов растений и что всем этим управляли процессы,
происходящие в недрах и на поверхности Земли. Наше
исследование истории развития животных, как и истории растений, основывается
на данных изучения ископаемых остатков. Но летопись ископаемых остатков
животных имея много пробелов, и пока не будут найдены руководящие ископаемые,
мы вынуждены заполнять эти пробелы умозрительным! рассуждениями. По мере того
как мы обнаруживаем и изучает все новые ископаемые остатки, некоторые из
построений предыдущих лет подтверждаются и становятся доказанными истинами,
другие оказываются неверными и заменяются положениями, более соответствующими
фактам. Развитие
растительного и животного мира Как
мы уже говорили, между животными и растениями существует коренное различие,
возникшее с самых первых шага развития жизни на Земле. Это различие
заключается в способе питания. Растение нуждается в основном в углекислом
газе и солнечном свете, с помощью которых оно путем фотосинтеза вырабатывает
органические вещества, служащие ему пищей и образующие ткани его тела.
Животные, как одноклеточные, так и многоклеточные, используют в пищу ткани
других организмов: или растений, или вторично других животных. Кроме того,
они дышат кислородом, а этот источник энергии также поставляется в атмосферу
и гидросферу растениями. Без пищ и кислорода
животные не могли бы существовать. В таком случае они, очевидно, не могли
возникнуть до тех пор, пока не появились растения и не начали в процессе фотосинтеза
выделять свободный кислород. Как
было показано в таблице 2, мы располагаем данными летописи ископаемых,
свидетельствующими о том, что в начале последнего миллиарда лет истории Земли
(до фанерозоя) и растения, и животные жили в море,
взаимно дополняя процессы жизнедеятельности друг друга. Растения высвобождали
кислород из углекислого газа, а животные дышали этим свободным кислородом,
окисляли растительную пищу и создавали углекислый газ, который снова
использовался растениями. Таково биохимическое взаимодействие растений и
животных. Однако,
хотя это взаимодействие продолжалось и позднее, животные начинали приобретать
все более и более отличный от растений облик. Растениям, жившим сначала в
мелководных морях, а затем на суше, не нужно было перемещаться в поисках
пищи, они могли просто поглощать ее, оставаясь на месте. На суше им для этого
требовались только листья, каждый из которых освещался солнцем, корневая
система для поглощения водных растворов из грунта и соединительное
устройство, служащее для поддержки листьев и подачи к ним растворов. Именно
такими приспособлениями обладает большинство растений. Животным же, за
исключением некоторых, наиболее простых, прикрепленных к морскому дну,
приходилось двигаться в поисках пищи, которой им служат растения и другие
животные. Животные - это охотники, и с самых первых шагов эволюции они
приобрели способность к передвижению. Наиболее древние животные, жившие в
море, изменялись различными способами. Путь самого успешного развития
представляет для нас наибольший интерес, так как в ходе его были заложены
основы строения человеческого тела. Это развитие состояло в образовании: во-первых,
прочного жесткого скелета, к которому прикреплялись мускулы; во-вторых,
хвоста, который использовался как рулевое весло; в-третьих,
системы конечностей (равного числа правых и левых), которые могут двигаться с
помощью мускулов, чтобы уравновешивать тело в воде и затем чтобы перемещать
его в толще воды; в-четвертых,
глаз, расположенных в передней части тела для того, чтобы видеть добычу и
препятствия, которые нужно избегать; в-пятых,
рта не только для того, чтобы поглощать пищу, но также и воду; в-шестых,
центрального управляющего устройства - мозга, соединенного с мускулами
нервной системой. Если
мы добавим к этому списку седьмой пункт - удлиненную, обтекаемую форму тела,
служащую для уменьшения трения при движении в толще воды, то становится
понятно, что получившийся в результате организм в основном подобен рыбе. Конечно,
не все эти усовершенствования появились сразу. Они развивались в течение
десятков, если не сотен миллионов лет. Ни одно из них не было чем-то
совершенно новым. Все они представляли собой дальнейшее развитие или
видоизменение уже существовавших свойств. Рыбообразная
форма тела была наиболее удачной для жизни в море. Позднее эта же конструкция
послужила основой для формирования тела наземных животных. Условия
обитания организмов в море Нетрудно
представить себе, что древнейшие животные должны были развиваться во
множество видов, потому что океан далеко не однообразен. В нем можно выделить
множество различных видов среды, в которых происходит (путем естественного
отбора) формирование организмов, характеризующихся многообразием форм и
специализаций. Миллиард лет назад в океане, как и сейчас, животные могли плавать
на поверхности воды или вблизи нее, могли населять холодное темное дно и
толщу воды между дном и поверхностью. На мелководье вблизи берегов, как и
сейчас, условия жизни различались в зависимости от температуры, вида коренных
пород или рыхлых отложений, слагающих дно, интенсивности перемешивания воды
волнами или течениями. Каждое даже самое небольшое отличие условий среды
оказывало влияние на организмы путем естественного отбора. Морские животные
приобретали специализацию в соответствии с условиями жизни. Некоторые из них
были растительноядными, поедавшими морские растения, другие - плотоядными,
питавшимися другими животными. Часть из них представляла собой так называемых
"мусорщиков" и поедала трупы животных. Другие были хищниками,
пожиравшими живую добычу, Несомненно, что уже тогда, в эти древние времена,
мелководные моря были оживленными местами; их многочисленные обитатели
занимались поисками пищи, спасались от хищников, поедали других животных или
сами становились добычей.
Фото 29. Реконструкция облика дна кембрийского моря.
Г - голотурии, М - медузы, ГБ - губки, Т - трилобиты. Размер трилобитов -
5-7,5 см Некоторое
представление о том, как выглядело дно мелкого моря в кембрийское время, дает
фото 29. Среди колышущихся морских водорослей находится колония ветвистых
губок, напоминающих капусту, голотурии, парашютовидные
медузы, а также вымершие ныне организмы, родственные современным
ракообразным, - трилобиты. Эта группа живых существ была очень точно
воссоздана благодаря тому, что их остатки хорошо сохранились. В одном из
районов Британской Колумбии, в самом сердце Скалистых гор Канады, один геолог
почти случайно нашел небольшие ископаемые остатки в слое черных сланцев.
Осторожно раскопав этот слой, ему удалось найти остатки беспозвоночных
среднего кембрийского периода, принадлежавшие к 130 видам; конечности и даже
усики животных сохранились до мельчайших деталей (фото 30); они были
превращены в уголь, как та рыба, которую мы видели на фото 15. Эта находка
представляется исключительной, так как обычно все съедобные части тел мертвых
животных на морском дне почти неизбежно уничтожаются трупоядными
организмами. В данном случае отложение происходило на дне моря, к которому
был прегражден доступ воды, содержащей кислород. В придонных слоях воды,
лишенных кислорода, не могли жить какие-либо организмы, кроме некоторых
бактерий. Находившиеся здесь организмы погибли, и их тела были погребены на
дне, в слое тонкого черного ила. Так как трупоядные
организмы тоже не могут существовать без кислорода, то эти тела сохранились
до мельчайших деталей, Бескислородные области
существуют в морях и океанах и теперь; одна из них находится в Черном море.
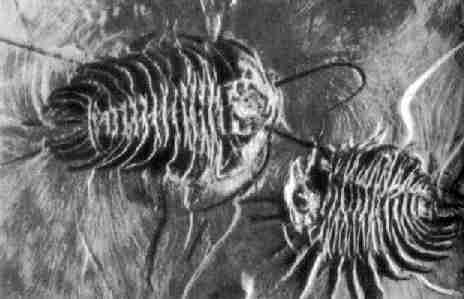
Фото 30. Два ископаемых трилобита (вида Olenoides seratus) в
кембрийских черных сланцах Британской Колумбии. Сохранились не только усики,
но даже длинные отростки на хвосте Среди
130 видов животных, извлеченных из этого черного сланца, совсем нет плеченогих моллюсков, несмотря на то, что около одной
трети кембрийских видов на всей Земле составляли плеченогие.
Отсутствие этих животных вряд ли было случайным, Мы вправе предположить, что
это связано с местными условиями среды. Плеченогие,
которые жили, прикрепляясь к грунту, не обладали подвижностью и не могли
плавать. Таким образом, они не могли жить на илистом дне, и, кроме того, они
были настолько тяжелы, что не могли быть занесены в эту мертвую зону
течениями. То
обстоятельство, что на этом небольшом участке морского дна создались условия,
благодаря которым сохранились многие ископаемые организмы, необычайно
обогатило наши знания о степени развития, достигнутой морскими
беспозвоночными в кембрийскую эпоху. Степень этого развития оказалась
довольно высокой. Видовой состав кембрийских беспозвоночных резко отличался
от современного. Остатки, собранные в кембрийских слоях, содержат в среднем
около 60% трилобитов, 30% плеченогих (фото 4) и
только 10% других видов. Сейчас трилобиты вымерли, плеченогие
обитают лишь на небольших участках океана, а преобладают именно "другие
виды". Участок
морского дна, изображенный на фото 29, может показаться перенаселенным. Но он
не более перенаселен, чем современные коралловые рифы, как может увидеть
каждый, кто ныряет с аквалангом. Отбор образцов и подсчет количества
ископаемых остатков с каждой единицы объема осадочных пород показывает, что
морское дно было повсюду густо населено, возможно, уже задолго до
кембрийского времени. Живые существа стараются заполнить все доступное
пространство, и если коренные обитатели вытесняются откуда-либо пришлыми
организмами, то последние затем также густо заселяют это пространство. Раковины и
скелеты Следует
отметить одну особенность обстановки, изображенной на фото 29, - присутствие
животных, обладающих раковинами и скелетами. Эти приспособления оказали
заметное влияние на ход эволюции. Губки имели кремниевые скелеты, а некоторые
микроскопические плавающие животные и растения, не показанные на этом
рисунке, обладали раковинами из того же материала. В панцирях трилобитов слои
фосфата кальция перемежались прослойками хитина - соединения азота,
обнаруженного также в ногтях человека; структура панциря напоминала лист
фанеры. Большинство многочисленных плеченогих
моллюсков и других животных, не изображенных на фото 29, обладали раковинами
из карбоната кальция. Раковины - это твердые части тела, которые обычно
сохраняются в ископаемом состоянии. Они представляют собой наружный скелет,
броню, которая выполняет полезную для животного функцию. Более ранние морские
животные имели мягкое тело, у них не было твердых частей, которые могли бы
сохраняться. И однако, здесь, в кембрийском море, более полумиллиарда лет назад
уже были животные, которые успели обзавестись раковинами трех различных
типов.
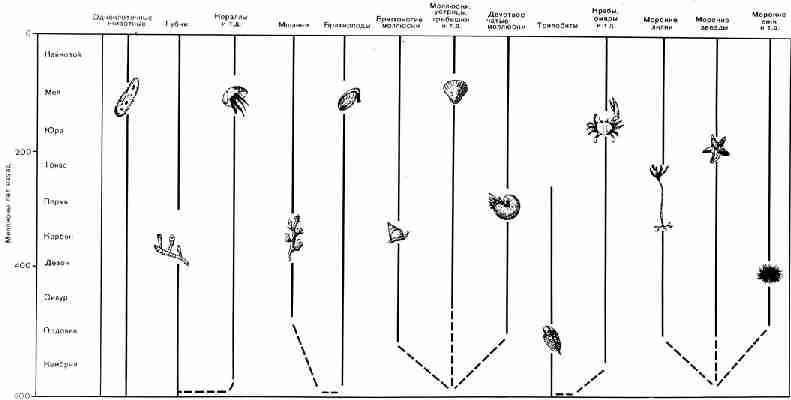
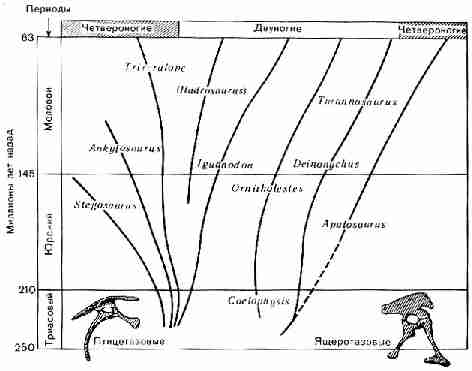
Рис. 40. Основные группы морских беспозвоночных и их
положение на геологической шкале. Пунктирными линиями показаны вероятные
эволюционные связи между некоторыми группами Палеонтологическая
летопись дает нам слишком мало данных, чтобы судить о том, как у морских
животных появились твердые части тела, и нам приходится полагаться на
теоретические рассуждения. Например, мы можем представить себе, что раковины,
состоящие из карбоната кальция, образовались следующим путем. Ионы кальция
присутствуют в морской воде и в большей части пищи морских животных. Поэтому
кальций мог проникать в тело животного. Большая часть его выводилась из
организма, но некоторое количество (зависящее от биохимических особенностей
животного) могло оставаться внутри тела или на его поверхности и
накапливаться, образуя своего рода затвердение. На морском дне, изобилующем
хищниками, вероятно, было безопаснее иметь броню, и эволюция способствовала
развитию этих затвердений. В результате постепенно образовались скелеты и
раковины. Так это происходило или иначе, но процесс отвердевания частей тела
морских животных шел параллельно с процессом развития сосудистой системы у
наземных растений. В
течение 60 миллионов лет, или около этого, кембрийского периода
беспозвоночные с наружным скелетом, имеющим форму раковины, были наиболее
сложным видом организмов. Высшие организмы с внутренним скелетом, каким
обладают позвоночные животные, в это время еще не появились. Краткий
обзор эволюции беспозвоночных Почти
все основные группы морских беспозвоночных присутствуют в виде ископаемых в
кембрийских слоях. Большая часть из них существует (в измененном виде) и в
наши дни. Все данные о развитии этих групп сведены воедино на рисунке 40, В
каждой группе происходили определенные эволюционные изменения, но в основном
эти изменения были не так резко выражены, как у наземных животных. В
большинстве случаев это определяется тем, что в море природные условия
менялись в более узких пределах, чем на суше. Тем не менее мы не должны
забывать, что корреляция пластов и составление геохронологической шкалы
(табл. 1) стали возможны главным образом благодаря наличию различающихся и
сходных черт, развивавшихся в процессе эволюции у морских беспозвоночных. Таким
образом, чтобы уделить больше внимания историк позвоночных животных, мы
ограничимся рассмотрением только двух примеров жизни на мелководном морском
дне в различные моменты времени, более поздние, чем изображенный на фото 29. Дно
моря в девонское время. Слои песчаников девонского возраста, выходящие на
поверхность в районе Гамильтона в центральной части штата Нью-Йорк, содержат
целую коллекцию ископаемых остатков. Образец этих пород показан на фото 31. В
центральной части его находится морская звезда Devonaster
euchans ("грациозная девонская звезда"),
которая очень похожа на современные виды морских звезд. В
"северо-восточном" углу образца находится o
ее более изящная родственница "хрупкая звезда" - офиура. В "северо-западном" углу, на внешнем
образца, находится цилиндрическое углубление окруженное кольцами. Это
отпечаток внешней поверхности стебля морской лилии, которая полностью изображена
в правой части рисунка 40. Большинство других ископаемых остатков принадлежат
трем или четырем видам плеченогих, группе животных,
имеющих раковины со створками, как у двустворчатых моллюсков, также
изображенных на рисунке 40.
Фото 31. Ископаемые остатки морских беспозвоночных
на плите среднедевонского песчаника из окрестностей города Гамильтона, штат Нью-Иорк. Описание см. в тексте. Реконструкция, на фото
32, основана на ископаемых ого возраста, найденных в данном районе Именно
по остаткам фауны этого типа была восстановлена картина, изображенная на фото
32. Ныряя с аквалангом в девонском море, вы могли бы на участке морского дна
около 60 сантиметров в поперечнике увидеть нечто подобное изображенному на
реконструкции; начальными буквами на рисунке даны названия всех изображенных
на нем животных. Особенно следует обратить внимание на двух представителей
головоногих одного с длинной прямой раковиной, а другого с волнистой,
закрученной. Оба принадлежали к той же самой группе хищников, что и
современные кальмары и осьминоги; все они имеют щупальца, снабженные
присосками, которые служат для того, чтобы хватать и удерживать добычу. Дно
моря в меловое время. На фото 33 представлен другой облик морского дна. Так
выглядела в меловом периоде поверхность юго-западной части современного штата
Теннесси. Головоногие с прямыми раковинами, которые
были широко распространены на более ранних этапах палеозоя, вымерли в ходе
эволюции. Вместо них появились моллюски с закрученными раковинами в форме
плоских дисков, напоминающих часовую пружину или моток клейкой ленты. В
широком устьевом отверстии располагалась голова моллюска, снабженная
щупальцами, захватывавшими небольших рыб и другую добычу. Другой особенностью
мезозойских морей (в отличие от палеозойских) было присутствие головоногих,
подобных кальмарам и носивших название белемнитов. Они имели внутреннюю
раковину, напоминавшую спинной хребет и состоявшую из карбоната кальция;
большинство этих раковин, сохранившихся в большом количестве в ископаемом
состоянии, напоминало толстый карандаш или тонкую сигару. Один из видов
белемнитов был гораздо крупнее остальных и имел длину более полутора метров.
Белемниты, как и палеозойские головоногие, быстро передвигались, выбрасывая
реактивную струю. В основном принцип их движения был тот же, что у
современных ракет, если не считать того, что выбрасывалась вода, а не газ.
Струя, выбрасываемая головоногими, представляла собой "изобретение"
способа движения, совершенно отличного от передвижения с помощью ног или
плавников. Кроме того, белемниты, подобно современным кальмарам, имели мешок
с чернильной жидкостью, которую они могли выбрасывать через воронку,
окрашивая позади себя воду, очевидно, для того, чтобы сбить с толку
преследователя. Эти особенности еще раз иллюстрируют своеобразие эволюции в
выработке приспособлений к окружающей среде.
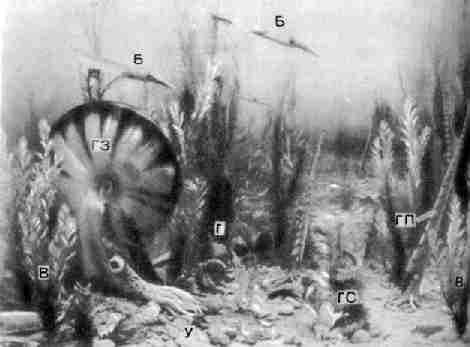
Фото 33. Картина дна мелового моря на месте
нынешнего города Мемфис, штат Теннесси. Ширина
изображенного участка дна около двух метров. В - водоросли; Б - белемниты; Г
- головоногие, с плоскозакрученной раковиной (ГЗ), как у современных наутилусов, с прямой раковиной (ГП) и со спиральной раковиной (ГС);
У - улитки "Морские
скорпионы". Особо следует упомянуть выделяющуюся своими исключительно
крупными размерами группу членистоногих (рис. 40), которые являются дальними родственниками
камчатских крабов и, возможно, предками современных скорпионов. Это
"морские скорпионы", которые жили в течение большей части палеозоя,
но достигли вершины своего развития в морях силурийского времени. Они дышали
жабрами, имели сегментную хитиновую оболочку, две пары глаз и шесть пар
членистых "ног" (фото 34). Некоторые их виды имели небольшие
размеры, но один был более двух метров в длину. Одни виды могли плавать,
другие ползали по илистому дну в устьях рек в поисках червей или небольших
рыб (фото 35). Превосходя по размерам современных им рыб, "морские
скорпионы" были самыми крупными животными, появившимися в биосфере в
этот период.
Фото 34. Углефицированный
ископаемый "морской скорпион" (Eusarcus scorpionis) длиной 10 см, найденный в силурийских породах
около города Буффало, штат Нью-Йорк. Ясно видны
членистое тело, ноги и хвост, кончик которого загнут вбок
Фото 35. Реконструкционный облик самого крупного из
известных "морских скорпионов" длиной более двух метров, а также
улиток и трилобитов на дне позднесилурийского эстуария около города Буффало, штат Нью-Йорк. На снимке справа, выше, показан
более мелкий скорпион, изображенный в виде ископаемого на фото 34 Насекомые.
Как и растения, беспозвоночные вышли из моря на сушу. Наземные моллюски
развились из морских моллюсков, насекомые и пауки развились из морских
членистоногих, однако об этом мы располагаем до сих пор лишь очень
отрывочными сведениями. Из-за малых размеров и отсутствия твердых частей в
телах этих животных они могли сохраниться в виде ископаемых лишь в совершенно
исключительных условиях. Таким образом, мы мало знаем о борьбе среди растений
и животных, переселившихся из моря на сушу. Ископаемые остатки позволяют
установить, что скорпионы, возможно, произошли от одного из более мелких
видов "морских скорпионов", существовавших еще в силуре, и что
немногие насекомые (пока еще бескрылые) и пауки существовали по крайней мере
с девона. В то время, как мы знаем, суша уже была покрыта примитивными
растениями. До некоторой степени организмы, переселившиеся на сушу, были подготовлены
естественным отбором для выработки дальнейших приспособлений к жизни на суше,
поскольку морские членистоногие уже имели защитную оболочку и обладали
способностью к активному перемещению. Главные изменения, которым они должны
были подвергнуться в процессе эволюции для того, чтобы окончательно
превратиться в насекомых, - это выработать систему органов дыхания и
научиться летать. Первую задачу они решили путем образования многочисленных
пор в оболочках; от пор отходили трубки, пронизывающие ткань самого тела.
Поэтому у насекомых дыхание и поглощение кислорода. осуществлялось всей
поверхностью тела. Вторую задачу насекомые решили к началу позднекаменноугольного времени путем образования выступов
оболочки тела, из которых развились крылья. В
таком сжатом виде эта история, пожалуй, выглядит повестью об удачливом
маленьком членистоногом, которое гналось за "успехами". Но прежде
чем нам начнет казаться, что здесь действовали целенаправленные усилия, мы
должны вспомнить о факторе времени. Промежуток от среднесилурийского
до начала позднекаменноугольного времени составил
приблизительно 120 миллионов лет - предположительно в десять раз меньше
времени потребовалось для образования Большого каньона реки Колорадо. За это
время сменилось по крайней мере 120 миллионов поколений членистоногих; этого
времени было вполне достаточно для выработки двух основных приспособлений
(для дыхания и полета), которые были "отобраны" из множества
попыток и экспериментов. Широко
распространенные в позднекаменноугольное время
угольные болота, на которых в мелкой воде происходило медленное накопление
глинистых отложений, представляли сочетание необычных условий, обеспечивавших
сохранность огромного количества насекомых в виде ископаемых. К настоящему
времени из них собрано и описано 800 видов только одних тараканов, один из
которых достигал 10 сантиметров в длину. Среди других ископаемых насекомых
попадаются виды длиной более 30 сантиметров и одна стрекоза с размахом
крыльев около 75 сантиметров, почти как у довольно крупного ястреба. Один из
этих великанов изображен на фото 26. Очевидно, мягкий климат,
благоприятствовавший пышной растительности каменноугольных болот,
способствовал также размножению насекомых. Не
удивительно, что эти насекомые, только недавно вышедшие из моря и пока еще
бескрылые, постепенно приобрели крылья и перешли к воздушному образу жизни.
Для мелких живых существ полет в воздухе давал наилучшие возможности
спасаться от врагов (от других насекомых, потому что птицы появились только
спустя 200 миллионов лет или даже позднее) и преследовать добычу. Необходимо
признать, что их образ действия оказался успешным, если под успехом понимать
способность заселять территорию и размножаться; доказательством этого служит
то обстоятельство, что сейчас число видов насекомых втрое превышает число
видов всех остальных групп. Большинство видов насекомых летающие. После
того как насекомые поднялись в воздух и приспособились к различным возможным
вариантам жизни в воздухе, потребность их в дальнейших изменениях стала
ослабевать и ход эволюции замедлился. В начале кайнозоя, после почти 250
миллионов лет развития, многие виды насекомых были уже очень близки к
современным. На фото 36 мы видим ископаемое насекомое, которое сохранилось в
тонкозернистых отложениях, образовавшихся на дне озера в Центральном Колорадо
около 30 миллионов лет назад. Это Glossina,
относящаяся к тому же роду, что и современная муха це-це
(переносчик сонной болезни), хотя и не к тому же виду. Таким образом, по
крайней мере у этого насекомого эволюция происходила довольно медленно.
Фото 36. Ископаемая Glossina
(муха це-це) из озерных олигоценовых отложений.
Рядом с ней - ископаемый лист дерева, родственного современным березам Происхождение
рыб Выше
мы описывали особенности формы и строения тела животных, подобных рыбам. И
все же остатков организмов, которые могли быть предшественниками рыб, не было
обнаружено. Наиболее обоснованное предположение (хотя это всего лишь предположение)
состоит в том, что объектом нашего поиска является предок современных морских
ежей и звезд, имевших продолговатое мягкое тело, лишенное скелета, не имевший
глаз, но обладавший бесчелюстным ртом для всасывания частиц пищи из
придонного ила. Одним из оснований для такого предположения послужил вид всех
остатков ископаемых рыб, принадлежащих к более ранней эпохе, - силуру (в
ордовикских слоях были найдены только костные пластинки). Остатки
свидетельствуют о том, что рыбы имели бесчелюстные сосущие рты и не имели
плавников. Их тела были одеты довольно гибким покровом из костяных чешуек, а
то, что их остатки были обнаружены в отложениях рек и озер, но не морей,
доказывает факт переселения этих примитивных рыб вслед за водорослями из
морей в реки и озера. На основании указанных фактов можно сделать два
заключения. Во-первых, удлиненное тело могло развиться, приспосабливаясь к
условиям постоянных речных потоков. Во-вторых, довольно грубые пластинки
костяной брони служили не для защиты от хищников (которыми в это время могли
быть "морские скорпионы"), а в качестве оболочки, препятствовавшей
избыточному проникновению пресной воды через поры внутрь тела. Такая
оболочка, вероятно, была необходима для животного, предки которого всегда
жили в соленой воде и у которого химизм тела был приспособлен к соленой воде. Таким
образом, девонские ископаемые включают морских рыб с челюстями, развившимися
из одной из жаберных дуг; они принадлежат к нескольким видам, и самой
замечательной из них была хищная рыба из рода Dinictis
("ужасная рыба"), у которой длина тела достигала 9 метров (фото
37). Акулы, изображенные на этом рисунке, обладали хрящевым скелетом, как и
современные акулы. Но у Dinictis строение несколько
другое, и в ее ископаемом скелете отмечается присутствие кальция. По пути
эволюции в этом направлении пошли костистые рыбы, которые также в девонское
время развились из более примитивных пресноводных предков. Их скелеты
полностью состояли из карбоната кальция и поэтому были более прочными и
жесткими. Плавники у них были приспособлены для более быстрого движения в
воде, увеличилась и обтекаемость тела. Такие преимущества обеспечили им
господствующее положение среди рыб, которое они сохраняют и до сих пор. Мы
можем считать их эволюцию успешной, потому что они дали большое количество
видов и стали более многочисленными, чем любая другая группа. Но так как
почти все эти рыбы продолжали обитать почти исключительно в море, то с точки
зрения общего хода эволюции они не получили в дальнейшем стимулов к развитию,
как это случилось с вышедшими на сушу животными.
Фото 37. Dinictis,
панцирная хищная рыба длиной девять метров, преследующая акул в мелководном
девонском море. Во рту у нее вместо зубов выступают заостренные зубчатые края
верхней и нижней челюстей Переход к
наземной жизни В
последней фразе мы сказали "почти все". Исключение составляла одна неприметная
группа костистых рыб, которая продолжала оставаться в пресных водах и, таким
образом, по существу, находилась на полпути к суше. В результате эти рыбы
имели гораздо более интересную историю развцтия,
чем их родственники, обитавшие в море. Им пришлось приспосабливаться к
изменяющимся условиям жизни на суше, они прошли такой же путь, который до них
прошли растения, а затем насекомые, и сумели "порвать" с водной,
даже пресноводной, средой и полностью выйти на сушу. Именно они стали предками
всех наземных позвоночных животных, в том числе, в конечном счете, и
человека. Эти
пресноводные представители группы костистых рыб послужили посредниками, через
которых человек получил костный скелет, надежно ему служащий. В своих
основных чертах скелет, подобный человеческому скелету, развился еще у
палеозойских рыб. Рыбы передали нам и основные черты нашего дыхательного
аппарата. Произошло это благодаря особенностям наших предков - пресноводных
рыб, одной из загадочных групп кистеперых рыб, потомками которых являются
современные кистеперые рыбы - целаканты (фото 10).
По другой линии развития они явились также предками современных двоякодышащих
рыб, которые в наше время обитают в реках сухих областей южного полушария;
они имеют вспомогательное легкое, развившееся из плавательного пузыря, с
помощью которого могут получать кислород из воздуха. С наступлением сухого
сезона двоякодышащие рыбы закапываются в ил на дне речки, образуя подобие
кокона, покрытого изнутри слизью. Свернувшись в этом коконе, рыбы впадают в
спячку. При этом скорость жизненных процессов составляет лишь 10% от
нормальной во время влажного сезона. При наступлении дождей ил размягчается,
рыба высвобождается из кокона и уплывает. Кистеперые
рыбы девонского времени имели легкие того же типа. Вначале следует отметить,
что в девонских речных отложениях были найдены скелеты Ichthyostega,
о чем мы уже упоминали в восьмой главе. Эта рыба принадлежала к амфибиям -
группе, название которой означает "ведущие двойную жизнь", то есть
частично на суше и частично в воде. К этой группе принадлежат современные
саламандры и лягушки. При сравнении амфибии Ichthyostega
с девонской кистеперой рыбой видно (рис. 41), что обе эти группы поразительно
близки между собой по размерам, форме и строению; особенно велико сходство в
строении плавников рыбы и ног амфибии. Еще раз посмотрев на рисунок 31, мы
можем провести такое же сравнение между девонской рыбой (кистеперой) и
изображенной рядом с ней амфибией. Очевидной, хотя и менее важной, чертой
сходства является то обстоятельство, что и у рыбы, и у амфибии голова покрыта
броней из костяных пластин. В целом сравнение не оставляет сомнений в том,
что девонские амфибии развились из девонских кистеперых рыб.
Рис. 41. Два этапа превращения рыб в наземных
позвоночных. А. Девонская кистеперая рыба с примитивным легким. Б. Девонская
амфибия Ichthyostega длиной около 90 см. Обратите
внимание на лапы с пальцами, все еще соединенными перепонкой, как плавники Безусловно,
эти перемены стали возможны благодаря добавочному легкому и особому
устройству плавника. Но что явилось толчком к появлению таких перемен? Для
объяснения были предложены две правдоподобные гипотезы. Согласно первой, основанной
на аналогии с современными двоякодышащими рыбами, причиной явилось изменение
климата и появление резко выраженных сухих сезонов, когда реки превращались в
разобщенные мелкие водоемы. Оставшись на сухом месте, кистеперая рыба была
вынуждена использовать свои необычные плавники для того, чтобы с их помощью
перетащить свое тело к ближайшему сохранившемуся водоему, в это время она
дышала своим примитивным легким. Экземпляры с менее эффективными плавниками
или легкими, или и тем и другим, не могли попасть в соседний водоем и
"отбраковывались". Выживали только обладатели наиболее эффективных
приспособлений. С течением времени некоторые из выживших рыб стали амфибиями. Другая
гипотеза предполагает, что причиной выхода кисте-перых
рыб на сушу явилось не высыхание водоемов, а стремление рыб спастись на
мелководье от хищников, которых было множество на более глубоких местах. Там
они использовали свои плавники сначала для поддержки тела, а затем и для
перемещения. Когда в результате естественного отбора плавники достаточно
окрепли, рыбы смогли выползать на соседние участки суши, где в девонское
время им еще не угрожали хищники и где могло завершиться превращение
плавников в примитивные ноги. Конечно,
обе эти гипотезы чрезмерно упрощают процесс, но в совокупности, видимо,
правильно отражают сущность происшедших перемен. Вспомним, что девонский
период длился добрых 50 миллионов лет - время, достаточное для того, чтобы
эволюция могла предпринять множество миллионов попыток. Можно было бы с
уверенностью предсказать, что за такое большое время какая-то группа озерных
или речных животных должна была выйти на прилежащие участки суши. Превращение
рыб в амфибий представляет собой заметное достижение на пути эволюции. Так
возникла новая династия - четвероногих позвоночных наземных животных. В
данной главе мы рассмотрели превращение мягкотелых морских животных в рыб,
которым свойственно удлиненное тело, поддерживаемое крепким скелетом,
состоящим из множества сочлененных костей, голова с глазами, рот, снабженный
челюстями и зубами, сильные плавники и хвост. Обладая всеми этими свойствами,
рыба должна была только усовершенствовать легкое, чтобы оно отвечало
требованиям среды. После этого процесса она могла полностью выйти на сушу. Глава тринадцатая. Амфибии и рептилии Амфибии Выход
амфибий на сушу. В предыдущей главе мы остановились на том, что первые
амфибии, только что развившиеся из кистеперых рыб, которые выползли на сушу,
стали знакомиться с сушей, которую им предстояло заселить. Конечно, обзор у
них был еще небольшой, так как на своих слабых и все еще очень коротких ногах
они были вынуждены ползать по земле, почти как рыбы, оказавшиеся на берегу.
Амфибии могли передвигаться только медленно и неуклюже, но по крайней мере на
суше у них не было врагов опаснее насекомых, которые, как можно предполагать,
их практически не тревожили. Но
даже при отсутствии опасности со стороны хищников или серьезных конкурентов
любые условия природной среды на суше представляют гораздо больше трудностей
для жизни ее обитателей, чем водная среда (в море, озере или реке). В море
условия жизни сравнительно однородны. Температура, освещенность и количество
пищи изменяются относительно мало. Поэтому морские позвоночные в условиях
стабильной среды изменялись меньше, чем наземные позвоночные. На суше животные
подвергаются непосредственному воздействию гораздо более многочисленных и
более действенных факторов, из которых достаточно назвать чередование тепла и
холода, палящего солнца и дождя, а также внезапные ураганы, наводнения и
пожары. В
этой новой среде позднедевонские амфибии, только недавно развившиеся из рыб,
были поставлены в такие же условия, как потерпевший кораблекрушение Робинзон
Крузо, неожиданно попавший в совершенно новую, незнакомую обстановку.
Робинзон нашел или, проявив немалую изобретательность, создал приспособления,
которые помогли ему справиться со всеми трудностями и выжить. Другими
словами, он приспособился к новой суровой жизни. Оказавшись
на суше, амфибии приспособились к некоторым свойствам своей новой среды.
Возможно, они "пошли" в определенной степени на компромисс,
ограничившись в своем расселении такими областями, где эти новые свойства
проявлялись слабее всего. С точки зрения истории эволюции их реакция была
совершенно нормальной. Организм остается на месте до тех пор, пока он не
вытесняется изменившимися условиями. Затем, если организм вообще способен
эволюционировать, то он изменяется ровно настолько, чтобы прийти в
соответствие с новыми условиями среды. Очевидно,
такая эволюция, содержащая элементы компромисса, дала возможность амфибиям
преодолеть опасности жизни на суше, поскольку, как можно судить по их
ископаемым остаткам, они продолжали существовать и образовывать новые виды в
течение всего палеозоя. По крайней мере 70 миллионов лет они были, бесспорно,
господствующими во всем мире наземными животными. Было найдено больше ста
видов ископаемых палеозойских амфибий. Такое большое число означает, что
животные приспособились ко многим условиям жизни вблизи воды (фото 38).
Некоторые из них жили вдоль берегов рек, как современные лягушки, другие - в
самой воде. Одни амфибии напоминали ящериц, другие - саламандр, а третьи
совсем не имели ног и походили на змей. Многие были похожи на короткохвостых
крокодилов, но это сходство было чисто внешним, так как крокодилы относятся к
рептилиям - более специализированной огруппе
позвоночных. Размеры амфибий колебались в больших пределах; длина их тела
составляла от нескольких сантиметров до трех метров. Многие из них, возможно,
имели чешуйчатую или гладкую кожу; другие, подобно своим двоякодышащим
предшественникам, имели на голове защитный покров из костяных пластинок.
Вероятно, в большинстве своем они были плотоядными и поедали насекомых,
червей или небольших рыб.
Фото 38. Четыре типа амфибий, живших в протоке с
чистой водой в одном из болот каменноугольного периода. Рисунок кожи амфибий,
который не сохранился у ископаемых, показан предположительно по аналогии с
ныне живущими амфибиями. Реконструкция Изменения
в процессе эволюции. Изучая многие виды ископаемых палеозойских амфибий, мы
можем выделить несколько основных изменений в анатомии амфибий. Эти изменения
произошли в течение первых десятков миллионов лет после выхода амфибий на
сушу. Небесполезно составить перечень этих изменений, так как некоторые из
них сыграли роль в истории развития человеческого скелета. 1.
Развитие шарового сустава конечностей. Плавник рыбы представляет собой
лопасть. Чтобы обеспечить движение в воде, ему достаточно колебаться только
вперед и назад относительно остального скелета. Но в наземных условиях
необходима также способность двигаться в стороны, а это требует большей
гибкости в бедренном и плечевом сочленениях конечностей. Такая необходимость
возникла благодаря окружающим условиям, и в результате естественного отбора
произошло развитие универсальных шаровых суставов. Эти суставы оказались
настолько полезными, что они были унаследованы от амфибий более поздними
наземными позвоночными и в результате стали непременной принадлежностью
человеческого скелета. 2.
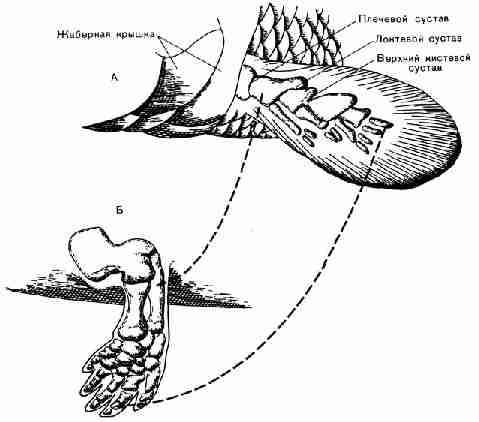
Развитие конечности с пятью пальцами. В плавнике кистеперой рыбы уже были
кости "запястья" и "кисти", но они имели продолжение в
виде тонкой перепонки, усиленной большим количеством тонких поддерживающих
костей - ребер; вся эта часть конечности напоминала часть конструкции зонтика
с натянутой тканью и спицами. По мере развития рыб "запястье",
"кисть" и их продолжение постепенно приобрели компактную
закругленную форму (рис. 42), число ребер уменьшилось до пяти, а перепонка
исчезла. В результате образовалась пятипалая конечность, очень полезное
приспособление для движения по неровной поверхности. Амфибии сохранили эту
особенность и передали ее своим потомкам.
Рис. 42. Превращение плавников рыб в пятипалую
конечность (не в масштабе). А. левый передний плавник девонской кистеперой
рыбы. Б. Левая передняя конечность палеозойской амфибии. (Пунктирными линиями
показаны соответствующие друг другу кости на рис. А и Б.) 3.
Удлинение конечностей. Когда кистеперые рыбы стали впервые использовать свои
плавники для опоры или придания телу устойчивости, они не могли приподняться
над поверхностью ила. Всякое движение у них носило характер ползания.
Напротив, амфибии, хотя их конечности все еще были короткими и
малоэффективными, могли удерживать свое тело, хотя бы какое-то время, приподнятым
над поверхностью земли, так что ползание превратилось в ковыляние. И это было
началом процесса ходьбы. 4.
Потеря панциря. Панцирь из костяных пластинок, который покрывал голову
кистеперых рыб, очевидно, был необходим для защиты от хищников в воде. Но после
выхода на сушу, где в те времена еще не было хищников, эта необходимость
отпала. Панцирь стал бесполезной и обременительной тяжестью, как солдатские
доспехи в Европе XVI в. после того, как вошло в
употребление огнестрельное оружие. В результате эволюционного процесса
панцирь исчез. Одна за другой группы животных лишались панциря, поскольку он
был уже не нужен в окружающих условиях. 5.
Увеличение размеров мозга. Процесс увеличения объема черепной коробки
(полости в черепе, содержащей мозг) ясно прослеживается по черепам ископаемых
амфибий. Но даже если бы такие ископаемые свидетельства отсутствовали, мы бы
все равно были уверены, что мозг амфибий должен был вскоре превзойти по
размерам мозг двоякодышащих рыб. Поскольку наземное животное подвергается в гораздо
большей мере воздействиям окружающей среды по сравнению с животными,
обитающими в воде, то условия суши в сильной степени вынуждают его развивать
свои умственные способности, с тем чтобы оно могло соответственно реагировать
на внешние факторы. Не обладая более совершенным механизмом реакций, амфибии
не смогли бы выжить. Упомянутые
пять изменений были важны сами по себе. Но даже вместе взятые, они еще не
могли скрыть очевидное сходство, существовавшее между ископаемыми рыбами и
амфибиями. Взгляните снова на рисунок 31, и вы увидите, насколько сильно это
сходство. Мы
уверены, что, помимо пяти изменений, прослеживающихся в ископаемых остатках,
за 50 миллионов (или около того) лет, которые протекли между позднедевонским
и позднекаменноугольным временем, амфибии
усовершенствовались еще в одном направлении, хотя по ископаемым остаткам
этого не видно. Должно было произойти развитие все еще довольно примитивного
легкого в более крупный и более развитый орган дыхания. При более быстром
передвижении, которое обеспечивали удлинившиеся ноги, требовалось больше
кислорода. Улучшение снабжения кислородом было также важно, если животное на
короткое время погружалось в воду, как это, должно быть, делала безногая
змеевидная амфибия, которую мы видим на фото 38. Несмотря
на исторический шаг, который сделали амфибии, будучи еще кистеперыми рыбами,
и в ходе которого они одержали двойную победу - начали дышать воздухом и
двигаться по суше, - несмотря на то что они первыми перешли к внутреннему
оплодотворению, дальше этого амфибии не продвинулись. Они не сумели перейти к
такому способу размножения, который позволил бы им порвать с водной средой.
Они навсегда остались привязанными к воде, потому что им приходилось
возвращаться к ней, чтобы откладывать яйца. Яйца современных амфибий, мягкие
и непрочные, как у рыб, должны откладываться в воде, где они могут быть
оплодотворены, и оставаться там, пока из них не выведется молодь. На воздухе
они быстро высыхают и не развиваются. После выхода из яиц молодь находится в
личиночной фазе (как, например, современные головастики) и дышит с помощью
жабр, как рыбы. Из-за
такой особенности процесса размножения амфибии занимают среди животных
приблизительно такое же положение, как бессеменные
в царстве растений. И тем и другим для размножения требуется вода. Поэтому
амфибии никогда не могли заселить возвышенные и сухие территории и по сию
пору стремятся оставаться в местах, где по крайней мере в некоторые сезоны
есть вода, в которую они могут откладывать яйца. Несмотря
на все эти несовершенства в строении амфибий, мы не можем так просто сбросить
их со счета, поскольку они занимают действительно важное место в эволюции
живых существ. Амфибии не только первыми из позвоночных прочно обосновались
на суше, но и внесли свой вклад в эволюционный процесс. Вклад этот состоял в
развитии таких особенностей строения, которые сделали возможным появление
рептилий, что в свою очередь вызвало новую волну усовершенствований. Подобно
тому как один из видов девонских рыб "выработал" легкое, так и один
из видов амфибий в позднекаменноугольное время
"усовершенствовал" строение яйца и тем самым положил начало линии
рептилий. Развитие такого яйца было большим достижением и стало еще одной
вехой на пути эволюции животных. Сейчас амфибии занимают далеко не
господствующее положение среди животных, но в течение по крайней мере 50
миллионов лет палеозоя они были доминирующей и развивающейся группой.
Пятьдесят миллионов лет - относительно продолжительный период; он больше, чем
тот позднейший отрезок времени, в течение которого все человеческие расы и
обезьяноподобные предки человека заселили Землю. Это время было достаточным
для многочисленных "проб и ошибок", которые привели к созданию
амниотического яйца. Рептилии Амниотическое
яйцо. Появление принципиально новой "конструкции" яйца (рис. 43)
ознаменовало новую эпоху, поскольку это яйцо не обязательно было откладывать
в водной среде. Оно было устроено таким образом, что имело запас воды внутри
себя, хотя и откладывалось на суше. Подобно космическому кораблю,
амниотическое яйцо содержало привычную "микросреду" и все
необходимое для жизнеобеспечения внутри своей защитной оболочки.
"Обитателем" яйца был эмбрион рептилии; таким образом, создав яйцо,
амфибии положили начало эволюционной линии рептилий. В настоящее время
подобного устройства яйца нет ни у одной амфибии, зато им обладают все
рептилии, а также все птицы и два вида млекопитающих. Конструкция яйца была
настолько удачной, что основные ее черты остались неизменными на протяжении
300 миллионов лет с момента его появления.
Рис. 43. Основные черты строения амниотического яйца Помимо
всего прочего, это яйцо имеет три оболочки, образующие три мешка. Первый, внутренний,
мешок - амнион (от которого яйцо получило название амниотического) - содержит
эмбрион, плавающий в жидкости. Жидкость и есть искусственная водная среда,
заменяющая озерную или речную воду, в которой амфибии и рыбы откладывали яйца
и в которой из яиц появлялись головастики, дышащие при помощи жабр, а позднее
превращавшиеся во взрослую особь, дышащую воздухом. Второй
мешок содержит желток, большой запас пищи для эмбриона. Третий - зародышевый
мочевой пузырь - связан с почками эмбриона и служит для сбора отходов - мочи
эмбриона. Все три мешка заключены в твердую, но проницаемую оболочку; изнутри
она выстлана другой оболочкой, позволяющей дышать находящемуся внутри яйца
эмбриону. Эта оболочка, играющая роль легкого, обеспечивает проникновение
внутрь яйца кислорода из окружающей среды и удаление из яйца углекислого
газа. Остальное пространство внутри яйца заполнено жидкостью, которая
разделяет все три мешка и играет роль гидравлического амортизатора,
защищающего эмбрион. Оплодотворение
амниотического яйца было внутренним и происходило внутри тела самки. Яйца рыб
оплодотворялись после того, как они отложены, и для того, чтобы выжить, рыбы
должны были откладывать много яиц. Как и голосеменные растения, они должны
были отличаться плодовитостью. Некоторые рыбы откладывают почти миллион
икринок в каждый сезон размножения. Икринок погибает так много, что
необходимо очень большое их количество, чтобы обеспечить выживание хотя бы
немногих. Внутреннее оплодотворение привело к гораздо большей надежности
размножения, потому что вероятность выживания отдельного оплодотворенного
яйца значительно возросла. Очевидно,
что появление амниотического яйца явилось еще одним звеном в цепи
усовершенствования способа размножения, начавшегося на ранних этапах
существования биосферы с появлением полового размножения растений.
Возникновение внутреннего оплодотворения и амниотического яйца было очень
важным этапом, но на нем усовершенствование не закончилось. Все
меры предосторожности, обеспечивающие безопасность беспомощного эмбриона,
покоящегося в своем мешке, как до, так и после откладывания яиц, выглядят
так, как будто бы они были предварительно продуманы и спланированы, но это
было не совсем так. По сути дела, это было ответной реакцией на воздействие
окружающей среды, осуществлявшейся через естественный отбор в каждом новом
поколении амфибий в течение очень долгого времени. Каким же именно было это
воздействие? Вполне возможно, что основное влияние оказало постоянное
увеличение в озерах и реках численности и разнообразия рыб и беспозвоночных
хищников, охотно поедавших икру и небольших личинок - головастиков,
беззащитную молодь амфибий. Создавшееся в результате этого положение не
представляло опасности для взрослых особей, но ставило под угрозу выживание
всего их сообщества. Появление любого приспособления, которое позволило бы
яйцам постоянно находиться вне воды и дало бы возможность молоди пройти эту
стадию личинок (когда они наиболее уязвимы) внутри яйца, а не в полной
опасностей внешней среде, было бы величайшим достижением. Сущность
происшедшего изменения можно сформулировать двояко. Мы могли бы сказать:
"Эта угроза, очевидно, заставила амфибий откладывать яйца в более
безопасных местах, там, где их не могли достать хищники, а именно на берегах
рек и озер выше уровня воды". С другой стороны, мы могли бы сделать хотя
и менее эффектное, но зато более точное предположение о том, что небольшое
преимущество с точки зрения возможности выживания имели те амфибии, у которых
генетические изменения шли в сторону увеличения прочности оболочки яйца,
содержавшего зачатки всех уже описанных нами составных частей. Дальнейшее
развитие и закрепление этих полезных изменений в ходе последовательной смены
поколений было только вопросом времени. А времени для этого было вполне
достаточно, как мы уже видели, рассматривая другие события в истории Земли.
От появления первых известных нам амфибий и до первых рептилий,
предположительно обладавших яйцом такого типа, прошло 50 миллионов лет. Амниотическое
яйцо неизбежно должно было возникнуть. Раньше или позже должно было появиться
или оно, или другое подобное устройство, которое привело бы к тем же самым
результатам. Поэтому вопрос состоял не в том, что именно должно появиться, а
в том, когда это произойдет. Хотя
в любой день мы можем позавтракать современным амниотическим яйцом, до сих
пор никто еще не находил ископаемых амниотических яиц в палеозойских слоях.
Наиболее ранние их находки относятся к слоям мезозойского возраста. Но позднекаменноугольные и пермские слои изобилуют скелетами
ископаемых рептилий, а все рептилии обладают яйцами амниотического типа.
Более того, у нас есть доказательства постепенного перехода от амфибий к
рептилиям, основанные на изучении не ископаемых яиц, а ископаемых скелетов. В
этих скелетах можно видеть в разнообразных сочетаниях признаки как амфибий,
так и рептилий. Все это, вместе взятое, позволяет установить факт перехода от
амфибий к рептилиям (фото 39). И если нас спросят: "Что появилось
раньше, рептилия или яйцо?" - то мы, основываясь на ископаемых
доказательствах постепенного перехода, должны будем ответить: "Ни то ни
другое; яйцо и рептилия развивались одновременно и постепенно, путем длинного
ряда небольших последовательных изменений".
Фото 39. Сеймурия (Seymouria),
наземное позвоночное животное из нижнепермских слоев, и его гнездо у подножия
дерева. Реконструкция, В скелете этого животного (длиной 60 см) присутствуют
в равной мере признаки рептилий и амфибий. Сначала предполагали, что это
рептилия, но теперь считают, что более логично относить его к амфибиям. Но
каково бы ни было положение сеймурии в классификации, это животное
представляет собой превосходный пример промежуточной стадии эволюционного
перехода одной группы в другую Только
что мы говорили о признаках амфибий и рептилий. Каково же принципиальное
различие между этими группами животных? Если начать с различий, заметных по
ископаемым скелетам, то мы должны обратить внимание на строение шеи и
конечностей. Рептилии имеют настоящую шею с одним шаровым суставом; таким
образом, их голова может свободно поворачиваться относительно позвоночника.
Животное, которое может, поворачивая голову, смотреть в разные стороны, может
быть более подвижным, быстрее ориентироваться как в поисках пищи, так и при
бегстве от врагов. У первых рептилий врагов, конечно, не было. Но прошло
немного времени, и враги появились из среды самих же рептилий. Рептилии
начали поедать друг друга; что-то подобное этому мы видим на фото 42. Что же касается
конечностей, то коротенькие и слабые конечности амфибий, развившиеся из
рыбьих плавников, в процессе перехода стали сильнее и удлинились. Таким
образом, они могли приподнять тело рептилий vнaд
землей и дали им возможность двигаться быстрее. Эти
различия и некоторые другие устанавливаются по ископаемым скелетам. Кроме
того, мы видим, что и мягкие части тел ныне живущих рептилий и амфибий
существенно различаются. Прежде всего вместо проницаемой кожи амфибий
рептилии обладают непроницаемым покровом, часто покрытым чешуйками или
пластинками жесткого рогового вещества. Очевидно, эти изменения
предназначались для того, чтобы уменьшить потерю воды на испарение через
кожу, но это могло произойти только после значительного усовершенствования
легких, потому что дыхание амфибий частично осуществлялось через их тонкую
кожу. Кроме
того, мышечная система и кровообращение рептилий более совершенны, чем у
амфибий; они имеют более развитое сердце-насос, обеспечивающий циркуляцию
крови. Наконец, мозг рептилий, хотя все еще небольшой по размеру, уже
обладает зачатками мозговых полушарий, этих крупных лобных долей мозга,
обусловливающих умственный уровень животного. Как известно, эта часть мозга
имеет большие размеры у млекопитающих и вообще отсутствует у амфибий; появление
ее у рептилий, возможно, объясняется улучшением кровообращения. Однако,
несмотря на все большие преимущества, рептилии не имели сколько-нибудь
эффективного способа регулирования температуры тела. Температура их тела
зависела от солнца: когда температура окружающей среды понижалась или
избыточно повышалась, рептилии становились вялыми и малоактивными. Последствия
возникновения амниотического яйца. Возникновение нового, более совершенного
яйца имело два следствия, очень важных для животных, которые обладали таким
яйцом. Наличие амниотического яйца повысило вероятность выживания, а также
дало возможность животным заселять новые более разнообразные типы территорий.
Стало выживать большее число зародышей, поскольку изменилось место откладки
яиц, а зародыши стали появляться из яиц уже более развитыми. Появилась
возможность откладки яиц за пределами досягаемости различных хищников,
уничтожавших яйца и молодь амфибий в воде. Кроме того, благодаря
приспособлениям, имевшимся внутри яйца, эмбрион мог полностью завершить свое
развитие и подготовиться к борьбе за существование к моменту выхода из яйца.
Значительно увеличилось также потенциальное жизненное пространство наземных
позвоночных. Причина этого, очевидно, заключалась в том, что рептилиям уже не
нужно было возвращаться к воде каждый раз, как наступал сезон размножения, и
они не должны были селиться вблизи водоемов. Ничто не препятствовало им
заселять более сухие районы, где прежде не ступала нога позвоночного и где до
тех пор у них не было конкурентов. В то время как рептилии расселялись,
амфибии остались привязанными к рекам и озерам, где большинство их обитает и
по сей день. Процесс отделения рептилий от их предков амфибий и разрыв с
привычными прибрежными условиями обитания напоминает отделение амфибий от их
предшественников - рыб и выход их из водной среды, в которой они раньше
обитали совместно. Завоевание суши позвоночными животными продвинулось еще на
один шаг. Климат
и эволюция рептилий. После того как амниотическое яйцо стало окончательно
служить признаком более прогрессивной формы жизни, появился новый фактор,
который, очевидно, способствовал дальнейшему развитию как самого яйца, так и
рептилий. Сравнительное изучение пермских и верхнекаменноугольных
слоев позволяет обнаружить между ними большие различия, указывающие на
изменение природной среды. Климат стал холоднее, и огромные болота
каменноугольного периода постепенно исчезли. Уровень поверхности, по крайней
мере некоторых континентов, повысился, и образовались многочисленные
высокогорные хребты. Причины таких изменений все еще не ясны, но, возможно,
это связано с взаимодействием движущихся плит коры и столкновениями
континентов. Но
какова бы ни была причина, следствием явилось то, что влажные районы -
естественная среда амфибий - стали встречаться реже. Тем самым был положен
конец расселению амфибий - процессу, который продолжался на протяжении всего
каменноугольного периода и которому способствовало наличие обширных
болотистых низменностей, расположенных почти на уровне моря. Для
существовавших тогда многочисленных и разнообразных амфибий не оставалось
места. Жизнь постепенно становилась трудной для животных, зависящих от воды.
Но для животных, обладавших амниотическим яйцом, это изменение климата не
было столь губительным. Только что появившиеся рептилии могли
довольствоваться многочисленными болотами и реками. Однако, не обладая
средствами для поддержания постоянной температуры тела, они были вынуждены
избегать более холодных областей. Ползающие
рептилии. После того как с возникновением амниотического яйца появились
рептилии, они полностью использовали преимущества, которые им предоставлял
этот способ размножения. Рептилии быстро изменили форму своего тела в
соответствии с разнообразными условиями среды обитания. Чтобы упростить наш
рассказ, мы можем опустить детали и выделить всего две группы -
"консервативную" группу ползающих рептилий и видоизмененную группу,
называемую динозаврами. Последние были настолько многочисленны и своеобразны,
что мы должны посвятить им отдельную главу, а сейчас мы кратко остановимся на
ползающих рептилиях. Говоря "ползающие", мы имеем в виду, что эти
рептилии продолжали традицию, унаследованную от амфибий, - их удлиненное тело
с короткими ногами находилось при движении у самой поверхности земли. По
форме тела ползающие рептилии в какой-то мере напоминали крокодилов (фото
40). Многие из них вели сходный с крокодилами образ жизни; очевидно, они
проводили много времени в реках и озерах, охотясь за рыбой. Развитие
позднепалеозойских ползающих рептилий было прослежено по ископаемым остаткам
в более молодых слоях; они постепенно разделились на две ветви, давшие начало
современным крокодилам и ящерицам.
Фото 40. Rutiodon, один из
многочисленных видов ползающих рептилий. Реконструкция. Наличие его
ископаемых остатков в нижнетриасовых слоях Европы и Северной Америки
позволяет предполагать, что в раннетриасовое время эти континенты, возможно,
соединялись. На переднем плане - цикадовые растения, позади рептилий - хвощи.
На заднем плане справа - папоротники, а за ними - хвойные растения.
Реконструкция Одна
из форм ползающих рептилий, ископаемые остатки которой в большом количестве
обнаружены в пермских слоях, имела характерное приспособление - бросающийся в
глаза парусообразный вырост на спине (фото 41). У
ископаемых скелетов этот "парус" состоит из длинных тонких костяных
отростков, которые отходили вверх от спинного хребта. У живой рептилии эти
отростки наверняка соединялись перепонкой, как пальцы утиной лапы. Измерения,
проведенные на многих ископаемых скелетах, показали, что площадь этого
выроста была пропорциональна объему тела рептилии. Очевидно, вырост был
пронизан кровеносными сосудами, и поэтому при нагревании солнечными лучами он
мог обеспечивать быстрое повышение температуры тела холоднокровной рептилии,
его действие можно сравнить с действием автомобильного радиатора,
охлаждающего воду, которая циркулирует по многочисленным трубкам. Первые,
хотя и несовершенные устройства такого типа появились у двух различных родов
ползающих рептилий. Один из них, диметродон,
изображенный на фото 41, был хищником, насколько мы можем судить по строению
его зубов. Но подобный же вырост появился и у совсем другой, травоядной
рептилии. У нее он отличался тем, что на длинных отростках находились
короткие поперечные выступы. Хотя это была несколько видоизмененная
конструкция выроста, его назначение было, очевидно, таким же, как и у
рептилий другого типа.
Фото 41. Диметродон (Dimetrodon), пермская ползающая рептилия длиной более трех
метров, имевшая на спине удивительное приспособление для терморегуляции. На
переднем плане хвощи; на заднем - папоротники и другие деревья. Реконструкция Рептилии,
подобные млекопитающим. Конец палеозойской эры ознаменовался быстрым
изменением рельефа и климата. Поэтому неудивительно, что в это же время
происходило и активное развитие рептилий, которые были тогда наиболее
развитыми и доминирующими животными. Среди вновь образовавшихся к этому
времени рептилий особенно важная роль принадлежит одной группе, потому что
именно она впоследствии дала начало новой эволюционной ветви, называемой
млекопитающими. Наиболее ранние свидетельства об этой группе содержатся в
ископаемых остатках из позднекаменноугольных и
пермских слоев, найденных в Южной Африке и Аргентине. Возможно, что эти две
области представляли в пермское время единый массив суши, как это показано (в
виде предположения) на рисунке 27. Если это так, то не удивительно, что на
этих двух континентах, в настоящее время разделенных тысячами километров морского
пространства, обнаружены ископаемые остатки одинаковых или очень схожих
наземных животных.
Фото 42. Два вида рептилий, подобных млекопитающим,
найденные в триасовых слоях Южной Африки. Три хищника (Cynognathus,
слева) готовятся напасть на травоядную рептилию (Kannemeyeria)
длиной около двух метров. Растительность представлена цикадовыми и хвойными.
Реконструкция Эти
ископаемые остатки представлены скелетами рептилий, но рептилий, у которых
уже начинают проявляться несомненные признаки млекопитающих; позднее в
триасовых слоях появляется несколько родов таких рептилий, и они еще в
большей степени похожи на млекопитающих. Отличительными их признаками,
прослеживающимися по скелетам, являются длинные шеи, дифференцированные зубы
и более развитый мозг. Предположительная реконструкция их облика дана на фото
42. Конечности удлинились, причем суставы на передних конечностях -
"локтевые" - сместились назад, а на задних - "коленные" -
вперед. Благодаря этому тело рептилии слегка приподнялось и стало не таким
приземистым. Подобное положение тела мы видим у млекопитающих (рис. 44), у
которых кости конечностей надежно поддерживают тело и освобождают мышцы от
излишней нагрузки. Зубы, которые у рептилий были все одинаковыми и имели
коническую форму, начинают подразделяться на резцы, клыки и коренные.
Увеличился и развился мозг, особенно его лобные доли.
Рис. 44. Положение тела рептилии и млекопитающего К
другим признакам млекопитающих, которые не устанавливаются по скелетам,
относятся: 1)
молочные железы для вскармливания детенышей; 2)
волосяной покров, защищающий тело от влияний внешней среды и препятствующий
потере тепла; 3)
теплая кровь; 4)
четырехкамерное сердце - наиболее совершенный насос, предназначенный для
снабжения мозга (и всего организма) кровью вне зависимости от колебаний
внешней температуры. Рептилии
после захода солнца становятся холодными и вялыми, млекопитающие же сохраняют
относительно высокую температуру тела. Возможно, жившие в триасе рептилии,
подобные млекопитающим, обладали большей частью этих признаков, по крайней
мере в зачаточной форме. Что касается размножения, то предполагается, что они
откладывали яйца так же, как и наиболее примитивные из современных
млекопитающих, но точно это не установлено. Однако мы можем с уверенностью
утверждать, что развитию более длинных конечностей сопутствует
усовершенствование мозга. Такие конечности позволяют уже не ползать, а
передвигаться с большой скоростью, что требует значительно лучшей мышечной
координации и вызывает необходимость в более сложном контролирующем
устройстве. Любой конструктор самолетов может сказать, что, чем большее число
выполняемых операций вы закладываете в конструкцию самолета, тем большим
числом контролирующих приборов вам придется его оснастить. Рептилии,
подобные млекопитающим, были многочисленны и разнообразны. Некоторые из них
превышали в длину три метра; другие были не больше мыши. И все они вымерли.
Но перед тем, как исчезнуть, одна из них - вероятно плотоядная, изображенная
на фото 42, - положила начало ветви настоящих млекопитающих, что явилось еще
одним крупнейшим достижением эволюции. Суммируя,
мы можем наметить четыре пути, по которым шло эволюционное развитие рептилий
(рис. 45). Первый - это наиболее консервативный путь ползающих рептилий,
которые никак не проявили себя и навсегда остались ползающими. Второй путь -
это путь динозавров, образовавших множество крайне своеобразных
разновидностей и тем не менее прекративших свое существование до начала
кайнозойской эры. Третий путь - это путь птиц, которые развились из рептилий
незадолго до начала юрского периода. Четвертый, и последний, путь начался с
рептилий, подобных млекопитающим. Он быстро привел к млекопитающим и в
кайнозое разделился на множество троп, одна из которых привела к человеку.
Рис. 45. Пути развития рептилий, ставшие возможными
благодаря появлению амниотического яйца На
этих путях эволюции происходило много неожиданного. Одной из наиболее
драматических была история динозавров, о которых мы расскажем в следующей
главе. Глава четырнадцатая. Век динозавров Рептилии
на суше За
палеозойской эрой последовал один из самых замечательных периодов в истории
жизни на Земле - царствование мезозойских рептилий. В пределах мезозоя, на
протяжении 190 миллионов лет, произошло поразительное распространение
рептилий. Рептилии, появившиеся в результате эволюции в позднекаменноугольное
время, благодаря преимуществам размножения с помощью амниотического яйца
распространились на суше, заселили моря и поднялись в воздух, используя
только что появившиеся крылья. Одна из ветвей рептилий дала начало птицам,
которые конкурировали с самими летающими рептилиями. Другая ветвь, как мы уже
видели, развилась в ветвь млекопитающих. И все же наиболее яркие персонажи в
драме рептилий - динозавры. Они и все их родичи, плавающие и летающие,
вымерли в мезозойскую эру. Вымерли полностью, до последней особи, предоставив
заселять Землю новым группам животных, главным образом потомкам мезозойских
млекопитающих. Летопись
ископаемых свидетельствует о существовании сотен родов динозавров размерами
от цыпленка до десятков метров, имевших десятки самых разнообразных
приспособлений к жизни в различных условиях. И все же, как и все другие
рептилии, все динозавры, вероятно, были холоднокровными и прямо или косвенно
зависели от наличия в большом количестве лиственной растительности. Однако
ископаемые динозавры были найдены на всех континентах, кроме Антарктиды. Из
этого обстоятельства мы можем заключить, что в мезозойское время были широко
распространены низменности с мягким климатом и пышной растительностью, так
как животные, подобные динозаврам, не могли существовать в высоких горах с
крутыми склонами и в холодном климате. Этот вывод, по-видимому, согласуется с
имеющимися у нас данными о перемещениях плит земной коры. Взглянув еще раз на
рисунок 27, мы увидим, что большая часть территории суши, которая сейчас
располагается в средних или высоких широтах, в середине мезозоя находилась
(предположительно) в более низких широтах. Не исключено, что южная часть
Северной Америки и южная часть Европы тогда примыкали к экватору. Если бы
континенты занимали в мезозое то же положение, что и сейчас, то маловероятно,
чтобы рептилии были столь многочисленны и чтобы достигали таких огромных
размеров. С
помощью карты, представленной на рисунке 38, мы можем объяснить расцвет
мезозойских рептилий с другой точки зрения. К концу мезозоя территория
Северной Америки по сравнению с настоящим временем в большей степени была
занята мелководными морями и в меньшей степени - сушей, в большинстве своем
низменной, а Мексиканский залив соединялся с Северным Ледовитым океаном. При
этих условиях климат центральной и северной частей континента, возможно, был
более мягким, чем сейчас, особенно зимой. И в Европе и на других континентах
в мезозое были распространены обширные моря. Таким
образом, расцвет рептилий в мезозойскую эпоху, который на первый взгляд
кажется необъяснимым, в конце концов удовлетворительно объясняется наличием
окружающих условий, которые были благоприятны для холоднокровных животных.
Таким образом, как и при рассмотрении более ранних этапов истории живых
существ, мы еще раз убеждаемся, что условия окружающей среды оказывают
определяющее влияние на развитие животного мира путем естественного отбора. Типы
динозавров Мы
уже говорили о том, что динозавров существовало несколько сот видов. Но все
они принадлежали к двум четко выраженным подразделениям, которые произошли от
одного общего предка в триасовое время, прежде чем появились динозавры как
таковые. Название "динозавр" является скорее популярным, чем
научным. Оно означает "страшный ящер", и когда его впервые ввели,
оно относилось к очень крупным и свирепым животным. Но динозавров такого типа
было относительно немного среди тех многочисленных рептилий, которых мы в
настоящее время относим к динозаврам. В эту группу входит большое число
рептилий, не обладавших ни свирепостью, ни большими размерами. Упоминавшиеся
основные два подразделения динозавров ученые различают на основании строения
их тазовых костей. К одному относились динозавры, у которых тазовые кости
имели такое же строение, как у ящериц, а ко второму - динозавры, тазовые
кости которых напоминали птичьи. Это важное конструктивное различие в
устройстве показано на рисунке 46. Нет необходимости останавливаться на этом
более подробно, нас интересует в первую очередь внешний облик и образ жизни
динозавров. Поэтому мы можем перейти к описанию отдельных наиболее выдающихся
представителей мира динозавров. Триасовые динозавры были довольно
примитивными и имели скромные размеры. Все они опирались на задние ноги, а
передние, гораздо меньшие по размерам, до земли не доставали (рис. 47). Шеи у
них были гораздо длиннее, чем у пермских ползающих рептилий. Однако, хотя
динозавры и стали двуногими, они не были прямо стоящими, как опирающийся на
две ноги человек. Когда они ходили или бегали, тело у них занимало положение
ближе к горизонтальному, чем к вертикальному, хотя, несомненно, они иногда
могли выпрямляться, как это зачастую делают белки. Что касается ног
динозавров, то, посмотрев на оставленные ими на влажном песке и иле следы
(фото 18), на которых отчетливо видны отпечатки трех или четырех длинных
пальцев и еще одного короткого, дополнительного, лишь изредка касавшегося
земли, мы можем понять, почему первые исследователи этих следов принимали их
за следы птиц.
Рис. 46. Связь между группами динозавров,
упомянутыми в книге Большинство
динозавров было хищниками, как и их пермские предки; присутствие среди
триасовых ископаемых разновидностей, обладавших необычного вида броней,
выростами и шипами, говорит о том, что они уже начинали "принимать"
защитные меры против своих врагов - других хищных динозавров.
Рис. 47. Coelophysis,
типичный динозавр триасового времени. Весьма вероятно, что мелкие следы,
показанные на фото 17, оставлены именно этим динозавром Естественно,
эта группа довольно примитивных триасовых динозавров включала предков всех
более поздних динозавров. Лучше всего подразделять их по способу питания,
образу жизни и особенностям строения. Мы можем выделить растительноядных и
плотоядных, двуногих и четвероногих динозавров, а также динозавров, имевших
броню, костяные пластины или защитные рога, и таких, кто не имел этих
приспособлений. Мы разделим рассматриваемых нами ящеров на четыре большие
группы. Растительноядные
двуногие. Хотя почти все раннемезозойские динозавры были хищниками, среди их
потомков встречалось много растительноядных особей. Судя по оставленным ими
следам, они довольно часто передвигались на четырех ногах. Среди них обычным
был игуанодон (рис. 48), животное плотного сложения, достигавшее около 11
метров в длину. В одном месте было найдено более 20 скелетов в полной
сохранности, по найденным совместно с ними скелетам черепах, крокодилов и рыб
можно думать, что эти динозавры жили в болотах. "Руки" у них имели
пять пальцев, причем "большой" палец представлял собой большой
острый шип, вероятно служивший неплохим орудием защиты. Очевидно, эти ящеры
питались, пригибая передними конечностями ветви деревьев и объедая побеги. Их
следы показывают, что они перемещались шагом и, вероятно, не очень быстро,
лишь временами делая короткие прыжки.
Рис. 48. Игуанодон, крупный двуногий травоядный
динозавр, обитавший в Европе Другая
группа растительноядных двуногих ящеров, достигавших 6-12 метров в длину и
носивших название гадрозавров, по образу жизни напоминала амфибий и обитала в
болотах или на их заболоченных берегах (фото 43). Между пальцами ног у них
имелись небольшие перепонки, а хвост был тонким, как у крокодилов, и при
движении в воде действовал, как весло. Ноздри были расположены так, чтобы
почти все тело могло быть погружено в воду. Рот состоял из рогового клюва,
подобного утиному. На челюсти находилось до тысячи зубов, длинных, очень
тонких, расположенных вплотную друг к другу. Когда роговой клюв доставал
мягкие растения из болота, верхняя и нижняя челюсти, на которых росли зубы,
начинали двигаться вперед и назад и тереться друг о друга, как две
проволочные щетки, перетирая таким образом пищу.
Фото 43. Гадрозавры (1), 'бронированный' динозавр,
подобный анкилозавру, (2) и плотоядный динозавр Struthiomimus
(3). Дерево слева относится к покрытосеменным. Реконструкция Плотоядные
двуногие. Там, где встречаются растительноядные животные, всегда есть и
хищники, которые на них охотятся. Среди динозавров было множество бегающих на
двух ногах хищников различного размера и облика. Один из них, Ornitholestes, длиной всего около двух метров, имел
настолько "изящное" строение, что весил предположительно менее 25
килограммов. Это было подвижное животное, приспособленное к быстрому бегу;
хватательные передние конечности с тремя очень длинными пальцами могли
поймать даже очень мелкую ящерицу, которая пыталась спастись бегством. Другой
динозавр, Struthiomimus (фото 43, под третьим
номером), был немного больше и напоминал страуса. У него даже был беззубый
клюв. Разбитый череп родственного ему динозавра был найден в ископаемом
гнезде, содержавшем яйца динозавров. Это обстоятельство, а также общий облик
животного, имевшего небольшой вес и гибкие "руки", приводит нас к
выводу о том, что Struthiomimus питался яйцами и
грабил гнезда. Еще
один динозавр, Deinonychus, длиной около 2,5
метров, который, возможно, был потомком динозавра Ornitholestes,
отличался двумя очень интересными приспособлениями, позволявшими ему вести хищный
образ жизни. Второй палец на каждой задней ноге был снабжен когтем, гораздо
более длинным и острым, чем все остальные когти. Этот палец имел особый
сустав, позволявший ему приподниматься над землей и поворачиваться на 180° (рис. 49), что позволяло рептилии наносить своей
добыче сильный удар ногой, удар, который мог распороть брюхо животному такого
же размера, как и сам хищник. Кроме того, в длинном хвосте этого динозавра
находились сухожилия, которые могли мгновенно "сращивать" кости
вместе, превращая хвост в жесткий противовес всему телу. Обладая подобными
когтями и хвостом, такой динозавр; должен был быть очень подвижным и опасным
животным.
Рис. 49. Deinonychus,
хищник, вооруженный острыми когтями Некоторые
двуногие хищники имели гораздо большие размеры, превосходя 9 метров в длину.
Один из них, тираннозавр, был самым большим из известных наземных хищников;
он имел длину до 15 метров, высоту до 6 метров и предположительно весил 7-8
тонн (фото 44). Длина его черепа составляла 1-2 метра, а во рту находилось
множество острых зазубренных зубов пятнадцатисантиметровой длины. Так как его
передние конечности были очень короткими, очевидно, он не пользовался ими,
когда нападал на добычу и поедал ее. Главной добычей тираннозавра были
растительноядные динозавры, как, например, гадрозавры и динозавры,
вооруженные рогами.
Фото 44. Тираннозавр, самый крупный хищник, нападает
на трицератопса, который приготовился к защите. Голова трицератопса покрыта
панцирным шлемом. Деревья представляют собой пальмы, относящиеся 'к
покрытосеменным растениям. Реконструкция Земноводные
четвероногие. Перейдем к гигантским динозаврам, которые так часто описываются
в популярной литературе, что их облик знаком не только ученым. В
палеонтологической летописи имеются данные по крайней мере о четырех
различных родах, внешне очень похожих; мы упомянем только двух из них. На
первый взгляд может показаться странным, что, хотя динозавры из этих двух
родов были четвероногими, передние ноги у них были гораздо короче задних. Но
в действительности этого можно было ожидать, потому что они были потомками
триасовых двуногих динозавров с короткими передними конечностями. Пожалуй,
лучше всего известен род Apatosaurus (фото 45) -
огромные малоподвижные травоядные, достигавшие 23 метров в длину; их короткое
туловище поддерживалось массивными колоннообразными ногами, снабженными
когтями. Впереди находилась длинная гибкая шея с маленькой головой, которую в
задней части туловища уравновешивал длинный гибкий хвост, сужающийся к концу.
Животное должно было весить более 30 тонн, то есть вчетверо или впятеро
больше самого большого африканского слона.
Фото 45. Apatosaurus,
четвероногий динозавр, подобный амфибиям, длиной более 20 м, на берегу
юрского водоема. Два других таких же динозавра пасутся в воде. По сравнению с
ними крокодил на переднем плане выглядит совсем маленьким. Растительность
состоит из цикадовых и хвощей. Реконструкция По
мере того как возрастали размеры и вес предков этого динозавра, происходила
эволюция скелета в сторону снижения его веса путем образования полостей и
отверстий в позвонках; тем самым вес снижался там, где нагрузки были
невелики, и сохранялся там, где была важна прочность, например в колонновидных ногах. Отпечаток ноги этого динозавра,
оставленный в мезозойских ил ах, превышает 90 сантиметров в длину. Другой
гигантский динозавр, диплодок, также был травоядным, во многих отношениях
подобным описанному выше. Основное отличие от него состояло в том, что
диплодок был несколько длиннее (длина одного экземпляра, согласно расчетам,
превосходила 29 метров при высоте почти 14 метров), но не так массивен, вес
его предположительно составлял 10-12 тонн. Без сомнения, большую часть
времени эти гиганты проводили в болотах и реках, поедая мягкие растения.
Вдали от берега, среди болотистых островов, они были в большей безопасности
от крупных хищников; поэтому такие места были для них не только
"столовой", но и убежищем. Для еще большей безопасности ноздри этих
гигантов помещались на самой верхней части головы, что позволяло им спокойно
дышать, почти полностью погрузившись в воду и находясь таким образом вне поля
зрения врагов. Эти и некоторые другие динозавры проглатывали свою
растительную пищу целиком и перетирали ее уже после того, как она попадала к
ним в желудок. Как цыплята, в зобу у которых обычно находится много камешков,
динозавры заглатывали камни размером с картофелину и с помощью этих орудий
своими сильными желудочными мышцами измельчали пищу. Иногда кучки таких
камней, когда-то окатанных и отшлифованных в желудках динозавров, находят
вместе с их скелетами, причем они располагаются там, где находилось брюхо
большого динозавра. Вероятно,
эти огромные животные откладывали яйца, хотя это не подтверждено еще
находками; яйца в воде погибали, поэтому они должны были откладывать их на
суше, а может быть на островах или в других местах, куда трудно было
проникнуть хищникам. Огромные
четвероногие динозавры, подобные амфибиям, обладали еще меньшим мозгом по
отношению к весу тела, чем другие динозавры, хотя и вся эта группа не слишком
славилась своими умственными способностями. У диплодока истинный мозг весил
только около семи граммов на тонну веса тела. Мы говорим "истинный мозг",
потому что диплодок, как и многие другие динозавры, имел дополнительный
координирующий центр гораздо большего размера, расположенный в позвоночнике,
вблизи таза. Этот центр соединялся с истинным мозгом через спинной и управлял
движением задних ног и хвоста. Хотя такое устройство может показаться
неудобным, мы должны признать, что оно действовало "исправно",
потому что им обладали многие различные виды динозавров, жившие на протяжении
десятков миллионов лет. Этому, конечно, способствовала среда обитания
динозавров с мягким климатом и мало изменявшимися природными условиями; в
подобной среде почти не возникало проблем, требующих умственного напряжения. Четвероногие,
снабженные броней или рогами. В наш перечень разнообразных динозавров входит
также довольно разнородная группа видов, которые, не будучи
близкородственными, обладали необычного вида броней, или рогами, или и тем и
другим. Несмотря на то что их предки в триасовое время были двуногими, эти
динозавры снова опустились на все четыре конечности. И все же их передние
ноги все еще были короче задних, как у Apatosaurus.
Будучи травоядными, они нуждались в защите от хищных рептилий; это и вызывало
развитие брони и защитных рогов. Наиболее
выделяющимся из этих бронированных рептилий был стегозавр. На его скелете,
имеющем в длину около 6 метров и вес предположительно 4 тонны, видны толстые
треугольные костные пластинки, окаймляющие спинной хребет, с которым они,
вероятно, соединялись связками. Возможно, эти пластинки, самые крупные из
которых достигали размера 75 сантиметров, защищали позвоночник от двуногих
хищников, которые, вероятно, при нападении пытались вцепиться в загривок, как
это делает терьер, убивая крысу. Кроме того, стегозавр был вооружен парой
крепких толстых шипов длиной около 60 сантиметров, расположенных на конце
хвоста. Один удар такого хвоста мог, вероятно, сбить с ног довольно крупного
противника и к тому же нанести ему серьезные повреждения. Анкилозавр
и его родственники (фото 43) имели, вероятно, такую защиту, какую имеют
современные броненосцы. Достигая 6 метров в длину и 2,5 метра в ширину, они
имели высоту менее 1,5 метра. Позади мощного, толстого, снабженного клювом
черепа вся верхняя половина тела у них была покрыта тяжелыми костными
пластинками. У некоторых из них вдоль всего тела, от плеч до хвоста,
напоминавшего тяжелую лопатку или дубинку, тянулись также огромные шипы.
Обладая такой защитной броней, эти рептилии двигались, вероятно, медленно. Но
при приближении опасности они могли прижиматься к земле, поджав под себя
лапы, и защищаться от нападения, нанося удары хвостом. Другим
способом, используя рога, защищался трицератопс и его многочисленные
родственники (фото 44). Эти громоздкие короткохвостые четвероногие достигали
7,5 метра в длину и трех метров в высоту. Их наиболее характерной
особенностью был огромный тяжелый череп, который протягивался назад в виде
большого щита, защищавшего шею. Спереди череп был снабжен двумя рогами,
выступавшими над узким клювом, похожим на клюв попугая. Внутри черепа
находился мозг, небольшой, по нашему мнению, но достаточно крупный для
динозавра. Наличие такого мозга позволяет предположить, что эти животные,
обладавшие защитным шлемом и рогами, были довольно подвижны. Об этом
свидетельствует незащищенность задней части их тела, не имевшей ни брони, ни какого-либо
оружия. Совершенно ясно, что они могли быстро поворачиваться, чтобы отразить
нападение врага рогами. Следами таких древних битв являются, возможно, шрамы,
часто встречающиеся на ископаемых остатках шейной брони. Рассуждая
о сражениях между динозаврами, мы невольно задаемся вопросом, происходили ли
они в тишине или сопровождались громкими воплями, как это происходит при
драках современных кошек и собак. Специалисты по анатомии динозавров могут
сообщить нам то немногое, что известно по этому вопросу. Представляется, что
конфигурация маленьких косточек у основания языка динозавров близка к той,
которую имеют те же косточки у некоторых ныне живущих видов животных.
Основываясь на этой аналогии, можно предполагать, что по крайней мере
некоторые динозавры могли издавать квакающие или лающие звуки, как это делают
современные крокодилы. Поэтому, если в палеозое на суше скорее всего царило
молчание, нарушавшееся лишь шумом ветра, потоков и прибоя, то мезозойские
ландшафты уже могли быть оживлены звуками, которые издавали животные. Протоцератопс,
родственный трицератопсу, но менее сложно организованный небольшой динозавр,
с клювом, но без рогов, живший в Азии, получил широкую известность в связи с
находкой его яиц и гнезд палеонтологической экспедицией в Монголии в
двадцатых годах нашего века. В позднем мезозое эта территория была такой же
засушливой, как и сейчас, и яйца были отложены в небольшие углубления в
песке, который в настоящее время превратился в песчаник. Самки динозавров
выкапывали ямки и откладывали в них до 15 яиц длиной 15-20 сантиметров. Было
найдено несколько таких гнезд, и по крайней мере в двух яйцах были обнаружены
крошечные кости детенышей динозавров, которые не смогли вылупиться. Были
найдены также яйца и других типов динозавров, как большего, так и меньшего
размеров. Морские
рептилии При
изучении жизни в мезозое едва ли не сильнее всего поражает то, что почти
половина всех известных видов рептилий жила не на суше, а в воде, в реках,
эстуариях и даже в море. Мы уже отмечали, что в мезозое на континентах широко
распространились мелководные моря, поэтому для водных животных не было
недостатка в жизненном пространстве. В
мезозойских слоях встречается большое количество ископаемых рептилий,
приспособленных для жизни в воде. Этот факт может означать только то, что
некоторые рептилии вернулись обратно в море, на родину, где когда-то давно
появились предки динозавров - рыбы. Этот факт требует некоторого объяснения,
поскольку на первый взгляд здесь происходил регресс. Но мы не можем считать
возвращение рептилий в море шагом назад с эволюционной точки зрения только на
том основании, что девонские рыбы вышли из моря на сушу и развились в
рептилий, пройдя стадию амфибий. Напротив, это положение иллюстрирует
принцип, согласно которому каждая активно развивающаяся группа организмов
стремится занять все разновидности среды, в которых она может существовать. В
действительности перемещение рептилий в море не слишком отличается от
заселения амфибиями рек и озер в позднекамен-ноугольное
время (фото 38). В воде имелась пища и конкуренция была не слишком жестокой,
поэтому сначала амфибии, а затем рептилии переселились в воду. Уже перед
концом палеозоя некоторые рептилии стали водными обитателями и начали
приспосабливаться к новому образу жизни. Эта адаптация шла в основном по пути
усовершенствования способа передвижения в водной среде. Конечно, рептилии
продолжали дышать воздухом так же, как дышит воздухом современный кит,
млекопитающее, хотя и сходное по форме тела с рыбой. Более того, мезозойские
морские рептилии развились не из какой-либо одной наземной рептилии, которая
приняла решение уйти обратно в воду. Ископаемые скелеты неоспоримо
свидетельствуют о том, что они имели различных предков и появились в
различное время. Тем самым ископаемые остатки показывают, насколько
разнообразным был отклик организмов на изменение условий среды, в результате
которого создалось обширное пространство, изобилующее пищей и пригодное для
заселения. Обширная
информация была получена при изучении ископаемых остатков, содержащихся в
морских аргиллитах и меловых известняках; в этих тонкообломочных породах
сохраняются не только кости, но также отпечатки кожи и чешуи. За исключением
наиболее мелких и примитивных видов, большинство морских рептилий было
хищниками и принадлежало к трем главный группам: их-тиозавры,
плезиозавры и мозазавры. Кратко характеризуя их, мы должны в первую очередь
отметить, что ихтиозавры приобрели сходную с рыбами удлиненную форму (рис.
50) и были превосходно приспособлены к быстрому плаванию в погоне за рыбой или
головоногими моллюсками. Эти животные, достигавшие 9 метров в длину, имели
голую кожу, спинной плавник и хвост, как у рыбы, а их четыре конечности
превратились в подобие тюленьих ласт и использовались для управления
движением тела при плавании. Все пальцы в этих ластах были тесно соединены, и
для увеличения прочности в них существовали дополнительные кости. Большие
глаза ихтиозавров были приспособлены для того, чтобы хорошо видеть в воде. У
них даже появилось одно очень существенное усовершенствование процесса
размножения. Будучи животными, которые дышали воздухом, но жили в морской
воде, они не могли откладывать яйца. Поэтому у ихтиозавров появился способ
размножения, при котором зародыш развивался внутри тела матери и, достигая
зрелости, рождался живым. Они стали живородящими. Этот факт устанавливается
находками превосходно сохранившихся остатков самок ихтиозавров с полностью
сформировавшимися детенышами внутри их тела, число детенышей достигает семи.
Рис. 50. Четыре группы животных, которые приобрели
обтекаемую форму тела в результате приспособления к жизни в воде: А.
рептилия, Б. рыба, В. птица, Г. млекопитающее. Первоначально они имели
различный облик, но в ходе эволюции приобрели внешнее сходство Вторая
группа включает плезиозавров, которые в отличие от рыбообразных ихтиозавров
сохранили исходную форму тела рептилий, достигая 7,5-12 метров в длину. Если
бы не хвост, плезиозавр походил бы на гигантского лебедя. Конечно, предком
плезиозавра была совсем не та наземная рептилия, которая дала начало
ихтиозаврам. Ноги у плезиозавров превратились в длинные плавники, а голова,
посаженная на длинной шее, была снабжена острыми зубами, которые смыкались и надежно
удерживали самую скользкую рыбу. Такие зубы исключали пережевывание;
плезиозавр заглатывал добычу целиком и затем измельчал ее в желудке с помощью
камешков. О рационе плезиозавров можно судить на примере содержимого желудка
одного из них, который, по-видимому, погиб прежде, чем камни в его желудке
успели измельчить проглоченную им пищу в должной степени. Было установлено,
что содержавшиеся в желудке кости и обломки раковин принадлежат рыбам,
летающим рептилиям и головоногим, которые проглатывались целиком, вместе с
раковиной. Третья
группа морских рептилий получила название мозазавров, потому что они впервые
были обнаружены вблизи реки Мозель, в северо-восточной Франции. Их можно было
бы назвать "запоздалыми", потому что они появились только в позднемеловое
время, когда ихтиозавры заселяли моря уже в продолжение почти 150 миллионов
лет. Предками мозазавров были скорее ящерицы, чем динозавры. Длина их
достигала 9 метров, они имели чешуйчатую кожу, а челюсти у них были устроены
таким образом, что они могли широко открывать рот, как змеи. Обтекаемое
тело как приспособление к условиям жизни в водной среде встречается не только
у ихтиозавров и мозазавров. То же самое можно видеть у целого ряда животных,
живших и раньше, и позже мезозоя, и в мезозое (рис. 50). Рептилии в
воздухе История
расцвета рептилий в мезозое не заканчивается тем, что изложено выше. Рептилии
не только распространились на суше и заполнили моря, они также поднялись в
воздух, следуя сразу по двум линиям эволюции. Они научились летать и как
рептилии, и кроме того, двигаясь совсем по другому пути развития, они
научились летать и как птицы. Насколько можно судить по ископаемым остаткам,
настоящие летающие рептилии были не такими многочисленными, как морские.
Однако они были первыми животными, поднявшимися в воздух после насекомых,
сделавших это еще в девонское время. Естественно, что воздушная среда труднее
для завоевания и более опасна, чем море. Для того чтобы двигаться в воздухе
или хотя бы пассивно парить, требуются более специализированные
приспособления, большая энергия и мастерство (под которым мы подразумеваем
подвижность и быструю реакцию), чем для передвижения в воде. В основном
именно поэтому человек построил корабли задолго до аэропланов. Промежуток
между этими изобретениями человека составил около нескольких тысяч лет. А
между возникновением рептилий в позднекаменноугольное
время и их проникновением в воздух (юрское время) прошло около 80 миллионов
лет. Мы
многое знаем о строении и облике летающих рептилий благодаря тому, что в
южной части Германии (ФРГ, Бавария. - Ред.) широко распространены осадочные
породы необычного типа. Эти породы представляют собой пласты известняка
позднеюрского возраста, настолько тонкозернистого, что он применялся для
гравировки иллюстраций для книг (до того, как для этой цели стали
использовать стальные и медные пластинки) и за это получил название
литографского камня. Необыкновенно тонкозернистый состав этих известняков
заставляет предполагать, что они отлагались в мелководных лагунах, защищенных
от волнения открытого моря песчаными барами или коралловыми рифами. Рыхлые
отложения на дне лагун сохраняли отпечатки даже мельчайших деталей растений
или тел животных, которые опускались на дно и заносились илом. В результате
литографский камень славится содержащимися в нем ископаемыми остатками
растений, беспозвоночных, рыб и рептилий.
Фото 46 Скелет Rhamphorhynchus,
примитивной летающей рептилии, найденный в литографских известняках в
Германии В
этих отложениях были найдены многие крылатые рептилии, кроме того, подобные
остатки были найдены и в других слоях мезозойского возраста в различных местах.
Рассматривая остатки одной из юрских примитивных рептилий, сохранившейся до
мельчайших подробностей (фото 46), мы видим, что ее тело приспособилось к
полету следующим образом: 1) уменьшился вес; 2) появились
"устройства" для управления полетом; 3) был создан механизм для
полета. Вот некоторые из этих приспособлений: 1.
Малые размеры тела; хотя некоторые летающие рептилии были с индейку, другие
были не больше канарейки. Скелет был облегчен благодаря развитию тонких полых
костей крыльев, а у некоторых видов и череп имел почти сетчатое строение и
состоял из тонких костей. 2.
Необычайно, хорошим развитием отличались глаза и часть мозга, управляющая
зрением. 3.
Наиболее замечательной особенностью были крылья. Глядя на рисунки 51 и 52, мы
можем легко себе представить, что четвертый палец на передней конечности,
словом "мизинец", - был необычайно удлинен по отношению к
остальным. От кончика этого пальца к задней ноге и далее к хвосту
протягивалась тонкая перепонка из кожи, образовывавшая крыло.
Рис. 51. Птеранодон (Pteranodon),
летающая рептилия с выростом на черепе; он пролетал огромные расстояния над
обширными меловыми морями на территории нынешних штатов Канзас и Небраска Все
эти три группы приспособлений, вместе взятые, создавали устройство, которое
хотя и неуклюже, но могло летать. Образование крыла, сопровождавшееся
усовершенствованием глаза и уменьшением общего веса, сделало возможным полет
привело к удивительным пропорциям тела. Например, одна из летающих рептилий с
размахом крыльев в 90 сантиметров, согласно расчетам, при жизни весила меньше
450 грамм. Кожа таких рептилий была голая, а челюсти снабжены многочисленными
острыми зубами, обычными для рептилий. Вероятно, эти животные, как
современные канюки, больше парили, чем летали. Происходя от наземных
хищников, они, очевидно, остались плотоядными и, медленно скользя над водой,
высматривали морских животных или крупных насекомых. Строение их скелета
показывает, что они не могли ходить. Очевидно, они приземлялись не на
поверхность земли, а на ветви деревьев или выступы скал на которых висели,
как современные летучие мыши.
Рис. 52. Схема сравнения крыла летающей рептилии,
летучей мыши и птицы. Все эти крылья появились в различное время. У рептилии
все крыло поддерживается только одним пальцем. У летучей мыши внешняя часть
крыла усилена четырьмя пальцами. У птицы большая часть крыла поддерживается
костями плеча и предплечья, а несущая поверхность образована легкими жесткими
перьями. Из всех трех видов такое крыло наиболее соответствует своему
назначению Позже
развитие летающих рептилий, остатки которых найдены в отложениях мелководных
морей мелового времени, пошло по пути замены зубов длинным клювом, который,
конечно, лучше соответствовал их образу жизни. У одного из родов в задней
части черепа появился особый выступ, или гребень (рис. 51), который,
возможно, уравновешивал длинный клюв и облегчал рептилии маневрирование при
ветре. Но главное изменение касалось увеличения площади крыльев, очевидно,
чтобы лучше поддерживать тело в воздухе. Одна из летающих рептилий для
поддержки тела, которое предположительно весило менее 12 килограммов, имела
крылья с размахом 7,5 метра. Такой размах крыльев позволяет считать этих
рептилий самыми крупными летающими животными за всю историю существования
жизни на Земле. Хотя до самого конца своего расцвета летающие рептилии
отличались хрупким сложением, все же они просуществовали более 100 миллионов
лет. Но
несмотря на то, что крыло рептилий выполняло свои функции и просуществовало
долго, оно было менее удачным приспособлением для полета, чем появившееся
независимо от него крыло птицы и позднее млекопитающих - летучих мышей. На
рисунке 52 показаны все три крыла, и, как можно видеть, крыло птицы из них
самое совершенное. Птицы В
юрское время у рептилий, живших по берегам теплых морей, существовали
различные виды полета. Мы уже видели, что несколько видов наземных рептилий
поднялись в воздух, используя для этого только что описанные кожистые крылья.
Но один из видов пошел еще дальше. В одном из карьеров при разработке
литографского камня в середине XIX в. был найден
ископаемый скелет рептилии, размером не больше вороны, имевшей большие глаза,
зубы, как у рептилий, и пальцы с когтями на передних конечностях.
Поразительно, что при этом были обнаружены очень четкие отпечатки перьев,
крепившихся к предплечью и к позвонкам длинного хвоста. Это, несомненно, была
птица. Она получила родовое имя археоптерикс (Archaeopteiyx)
("древнее крыло") и видовое название Uthographica
по названию горной породы (фото 47). В том же самом слое были обнаружены еще
два ископаемых скелета и отдельный отпечаток пера.
Фото 47. Археоптерикс (Archaeopteryx),
древнейшая известная нам птица, сидит на ветке хвойного дерева, собираясь съесть
пойманную ящерицу. На переднем плане справа - цикадовые растения; сзади -
хвойные деревья и еще одна такая же птица. Реконструкция Конечно,
эти находки представляли необычайный интерес и потому были тщательно изучены.
Результаты исследований могут быть, очевидно, сведены к следующему:
археоптерикс по своим основным чертам - летающая рептилия, но так как, по
определению, птицы имеют перья, а рептилии - нет, то ее можно отнести к
птицам. Особенности строения археоптерикса позволяют уверенно сказать, что эта
древнейшая из известных нам птиц произошла от жившей на земле двуногой
рептилии. Наличие перьев убедительно свидетельствует о том, что она имела
теплую кровь, потому что одной из основных функций перьев является
термоизоляция. У многих птиц кровь даже теплее, чем у человека. Их перьевой
покров и высокая двигательная активность позволяют им поддерживать нормальную
температуру тела около 39,5°С. Перья
состоят из того же прочного рогового вещества, что и чешуя. Некоторые ученые
предполагают, что небольшая рептилия, которая была предком этих примитивных
птиц, имела чешую и что сначала чешуйки стали волнистыми по краям, возможно,
потому, что такая форма предохраняла кожу от перегрева солнечными лучами.
Волнистые края оказались полезны и в другом отношении, поскольку они
уменьшали потерю телом тепла, и постепенно эти чешуйки превратились в перья.
Жесткость и небольшой вес вещества перьев сделал их идеальным приспособлением
для полета. Хотя
первая птица и имела перья, она, как и ее родственники - летающие рептилии с
кожистыми крыльями, летала плохо. Ее строение указывает на то, что птица,
вероятно, была хорошо приспособлена к планирующему полету. Возможно, она жила
на суше и, будучи хищником, питалась или мелкими животными, или падалью. То,
что ее остатки были найдены в морских известняках, свидетельствует лишь о
том, что отдельные экземпляры были унесены в море ветром или течением и
погребены в мягком донном иле. Хрупкие тела птиц, погибших на суше, просто не
сохранились. К
меловому периоду неуклюжесть полета у птиц исчезла и многие из них вместо
зубов обзавелись клювом. Некоторые птицы приспособились к жизни на воде.
Примером является весьма походившая на гагару плавающая и ныряющая птица гесперорнис (Hesperornis) (рис.
50), длина которой была около двух метров и которая все еще обладала зубами и
крыльями, хотя и не такими сильными и меньшего размера, чем у летающих птиц.
Существование птиц, которые почти покинули воздушную среду, перейдя к
плаванию, свидетельствует о том, что на ранних этапах развития птицы охотились
на рыбу так же, как это постоянно делали рептилии с начала мезозойской эры. Конец
огромных рептилий Окончание
мелового периода, означавшее конец всей мезозойской эры, можно назвать
"кризисом" в истории биосферы, потому что в это время произошло
вымирание многих групп животных. Наиболее заметный урон понесли рептилии.
Вымерли все динозавры, все летающие рептилии и все морские рептилии, за
исключением морских черепах; выжили и продолжили линию рептилий только
ящерицы, змеи и черепахи. Из беспозвоночных исчезли большинство головоногих,
в том числе все белемниты, а также некоторые линии морских двустворчатых
моллюсков и улиток. Вымирание
носило выборочный характер, потому что млекопитающие и наземные растения были
им мало затронуты или совсем не затронуты, а рыбы и многие беспозвоночные
полностью уцелели. Поэтому попытки приписать это вымирание какой-либо одной
причине оказались неудачными. Пока время основных событий в истории Земли не
было установлено с помощью радиометрического датирования, конец мезозоя
обычно именовался временем "великого вымирания". Однако сейчас мы
сознаем, что это выражение не соответствует истине. По крайней мере два
обстоятельства свидетельствуют о том, что вымирание не носило характера
катастрофы, уничтожившей все живое. Во-первых,
оно было избирательным, затрагивая одни виды и щадя другие. Кроме того, оно
не было ограничено каким-либо одним видом природной среды, охватывая сушу,
море и воздух. Во-вторых, хотя наиболее заметным было вымирание видов в конце
мелового периода, оно заняло в целом значительный промежуток времени. В
частности, различные группы рептилий вымирали в отдельные моменты времени в
течение всего мезозоя. Поэтому, какова бы ни была причина этого явления,
очевидно, она не вызвала "внезапного" истребления видов, по крайней
мере в том смысле этого слова, в каком мы применяем его к событиям истории
человеческого общества. Даже наиболее ярко выраженное вымирание,
происходившее в конце мелового периода, вероятно, тянулось несколько
миллионов лет. Рассматривая
геологические данные, говорящие о том, что происходило в конце мелового
периода, мы видим, что континенты в целом стали выше. В то же время и,
возможно, главным образом в результате этого поднятия сократились площади
обширных мелководных морей на континентах и исчезли болотистые низменности,
располагавшиеся по берегам этих морей. Кроме того, понизилась температура,
частично благодаря поднятию и сокращению площади морей. Мы
должны признать, что действительная причина вымирания до сих пор не
установлена. Ранее выдвигавшиеся объяснения - болезнь, недостаток пищи и
самое туманное - "потеря жизненной силы" - совершенно не в
состоянии объяснить, почему произошло выборочное исчезновение некоторых из
обитателей суши, моря и воздуха, а не полное вымирание обитателей какой-либо
одной среды. Представляется, что млекопитающие вышли из этой катастрофы
целыми и невредимыми. Недавно
было выдвинуто предположение, что конец мезозоя ознаменовался серией
обращений магнитного поля Земли (описанных в шестой главе) и что эти
обращения могли каким-либо образом повлиять на биосферу, например путем
изменения интенсивности радиации, поступающей на земную поверхность. Против
этого выдвигались возражения, но, возможно, сейчас еще слишком рано оценивать
все "за" и все "против". Достаточно сказать, что
вымирание, которым ознаменовался конец "века динозавров", до сих
пор представляет собой одну из величайших загадок, связанных с историей жизни
на Земле. Глава пятнадцатая. Млекопитающие Как
было сказано выше, шкала геологического летосчисления была построена на
основе ископаемых остатков. Фанерозой, часть этой
шкалы, охарактеризованная многочисленными ископаемыми, которые легко было
обнаружить, логически подразделяется на три крупных раздела: палеозой
("эра древней жизни"), мезозой ("эра средней жизни") и
кайнозой ("эра новой жизни"). Слово "жизнь" в каждом из
этих названий подчеркивает тот факт, что эти подразделения основаны на
содержащихся в горных породах ископаемых остатках организмов. В течение
кайнозоя господствующими были млекопитающие, мезозоя - рептилии. Основой
такого различия является изменение природных условий на континентах и в
морях, вызванное главным образом расширением морского дна и движениями
континентов. Северная Америка в кайнозойскую эру очень сильно отличалась от
того континента, который существовал в мезозое, а изменение условий жизни
вызвало эволюцию и возникновение новых форм жизни. Историю млекопитающих
лучше всего начать с рассмотрения современных нам млекопитающих, чтобы лучше
понять пройденный ими путь эволюции, который привел их к современному
состоянию. Мы
уже говорили, что характерными особенностями млекопитающих являются молочные
железы для вскармливания детенышей, теплая кровь (что фактически означает
постоянную температуру тела днем и ночью), волосяной покров, служащий для
термоизоляции, четырехкамерное сердце, дифференцированные зубы, служащие для
откусывания и жевания, и хорошо развитый мозг. Эти черты общие для всех
млекопитающих. По особенностям же процесса размножения млекопитающие
подразделяются на три группы. 1.
Наиболее примитивные - однопроходные, потому что они откладывают яйца (хотя и
вскармливают детенышей молоком); они отличаются также некоторыми
особенностями анатомии, которые в ископаемых остатках не устанавливаются.
Сейчас их существует только два рода - утконос с клювом, как у утки, и
австралийская ехидна (Отряд однопроходные объединяет два семейства: Tachyglossidae (ехидны) и Ornithorhynchidae
(утконосы). В составе семейства ехидны - два рода, утконосов - один род. -
Прим. ред.). Оба эти рода очень хорошо приспособлены к определенному образу
жизни и ограничены в распространении Австралией, где, по-видимому,
конкуренция слабее, чем в других частях света, что и создает предпосылки для
выживания самых примитивных животных. Поскольку ископаемые однопроходные встречаются
только в Австралии, возможно, что в других местах они никогда не
существовали. 2.
Промежуточную группу образуют сумчатые, из которых, пожалуй, наиболее
общеизвестны кенгуру. Детеныши у кенгуру рождаются живыми, они очень слабо
развиты; новорожденный кенгуру гложет иметь длину до трех сантиметров.
Детеныши заползают в сумку матери и в течение долгого времени питаются там
молоком, пока не вырастут в несколько раз больше (по сравнению с размером,
который они имели при рождении) и смогут выйти из сумки и кормиться
самостоятельно. Сейчас сумчатые живут главным образом в Австралии; к ним
относятся кенгуру, "медведи" коала и вомбаты. Единственное
сумчатое, сохранившееся в Северной Америке, - опоссум, - по-видимому,
приспособилось к конкуренции, живя на деревьях. Размножение у сумчатых более
безопасно, чем откладывание яиц, потому что развивающиеся детеныш постоянно
находится под защитой матери. Но еще более усовершенствовала этот процесс
третья группа млекопитающих. 3.
Наиболее развиты плацентарные млекопитающие, названные так потому, что у них
есть плацента - орган, находящийся внутри матки и напоминающий мембрану,
пронизанную многочисленными кровеносными сосудами. Через плаценту эмбрион
получает питательные вещества и кислород, и через нее же происходит удаление
углекислого газа и отходов. Все это устройство (рис. 53) напоминает
амниотическое яйцо (рис. 43), от которого оно, очевидно, и произошло. Но
плацента дает млекопитающим преимущества не только по сравнению с
яйцекладущими животными, но и по сравнению с сумчатыми. Эмбрион, развитие
которого регулируется плацентой, может находиться в матке до своего полного и
окончательного развития. Детеныш плацентарных млекопитающих, по крайней мере
у большинства родов, после того, как он родится, довольно скоро может
заботиться сам о себе. Плацента внесла существенное усовершенствование в
процесс размножения.
Рис. 53. Основные черты зародыша и матки у плацентарных
млекопитающих (сравните с рис. 43). На ранних стадиях зародыш питается
желтком, который затем исчезает. Зародыш получает питательные вещества и
кислород и выделяет углекислый газ и отходы через плаценту. Направление этого
обмена показано стрелками Мезозойские
млекопитающие С
нашей "позднекайнозойской" точки зрения,
современные млекопитающие находятся почти в конце эволюционной линии.
Рептилии пермского и триасового периодов, которые походили на млекопитающих,
находились на другом конце эволюционной линии. Необходимо помнить, что путь
развития организмов от наиболее примитивных млекопитающих до человека не был
прямым и это развитие происходило не всегда с одинаковой скоростью. В течение
почти 250 миллионов лет мезозоя эволюция млекопитающих не представляла ничего
интересного. Можно сказать, что они выжидали своего часа. В то же самое время
настоящие рептилии распространились очень широко и приспособились к различным
условиям среды в мягком климате. Низменный рельеф и мягкий климат мезозоя
способствовали развитию рептилий, но не представляли исключительно
благоприятных условий для млекопитающих. Вновь появившиеся млекопитающие
должны были выжидать, пока не уменьшились численно и не вымерли динозавры и
пока в кайнозое не возникли новые типы природной среды, лишь после чего они в
свою очередь смогли сильно распространиться и занять господствующее
положение. Все это ясно говорит нам, что эволюционное развитие меняет свое
направление в соответствии с меняющимися возможностями, которые предоставляет
окружающая среда. Пока происходит движение плит коры и перемещение
континентов в более высокие или более низкие широты, а иногда и соединение
континентов, будет происходить и изменение условий жизни, создающее новые
возможности для эволюции живых организмов. Вполне
вероятно, что предком настоящих млекопитающих мог быть Cynognathus
(фото 42) или один из родственных ему видов. А может быть, эта честь
принадлежала другой рептилии, еще не обнаруженной в ископаемом состоянии. Это
не имеет большого значения, потому что млекопитающие и рептилии, подобные
млекопитающим, очень тесно связаны между собой. Какой бы ни была рептилия -
родоначальник млекопитающих, - произошедшие от нее млекопитающие имели очень
небольшие размеры. Большинство ископаемых млекопитающих, обнаруженных в
триасовых, юрских и меловых слоях, были размером с крысу или мышь, а самые
крупные не больше кошки. До сих пор не найдено ни одного целого черепа, не
говоря уже о скелете, но обнаружено множество мелких челюстей с
дифференцированными, как у млекопитающих, зубами и разрозненные кости. Эти
кости больше напоминают кости сумчатых или однопроходных, чем плацентарных
млекопитающих. В меловых слоях в западной Канаде найдены остатки,
принадлежащие, без сомнения, опоссуму. Небольшие
размеры мезозойских млекопитающих указывают на то, что они не могли
конкурировать с динозаврами и другими рептилиями, несмотря на имеющийся у них
сравнительно большой мозг. Можно предполагать, что млекопитающие занимали
подчиненное положение, старались держаться в стороне и, вероятно, питались
насекомыми и семенами растений. Так как мы не располагаем целым скелетом, мы
не можем представить себе, как они выглядели, но ископаемые черепа позволяют
воссоздать внешний вид головы этих млекопитающих (рис. 54).
Рис. 54. Голова позднемезозойского млекопитающего.
Остальные части тела реконструировать пока не удается, так как найден только
череп Приспособления
к кайнозойским природным условиям Общий
обзор. Таково было положение вещей в конце мезозоя. На сцене начали
появляться млекопитающие. С изменением ландшафтов и похолоданием климата
динозавры постепенно исчезали, одна группа за другой, оставляя жизненное пространство
свободным. Млекопитающие, которым больше не угрожали рептилии, осмелели.
Волосяной покров и теплая кровь, а также больший объем мозга давали им
возможность выжить в условиях похолодания и приспособиться к новым, более
разнообразным природным условиям, на что динозавры оказались неспособны. По
мере того как рептилии сдавали свои позиции, млекопитающие занимали
освободившиеся территории, где им не угрожала существенная конкуренция.
Наконец рептилии исчезли. Единственной опасностью для этих первых млекопитающих
были другие млекопитающие. Конечно, среди "других млекопитающих"
вскоре появились плотоядные хищники, которые завоевали свое место в пищевой
цепи. В
этой истории можно найти сходство с мезозойской историей рептилий. Новая
форма распространяется на новые территории и там, встречаясь с различными
условиями среды, которую можно заселить, разделяется на несколько
разновидностей. Животные одного вида охотятся на других. В результате
конкуренции устанавливается своего рода динамическое равновесие видов. Оно
"динамично" потому, что при этом одни виды вымирают, другие
развиваются. Это равновесие представляет собой бесконечный ряд
"битв", в которых перевес получает то одна, то другая сторона. Если
кайнозойскую историю млекопитающих рассматривать в перспективе, она состоит
как бы из двух фаз: ранняя фаза экспериментирования природы на млекопитающих
и поздняя фаза отработки их деталей, во время которой направления эволюции
становятся более отчетливо выраженными. Ранняя фаза продолжалась включительно
по олигоцен, поздняя - остальную часть кайнозоя. Конечно, она продолжается и
сейчас, но с тех пор, как в конце плиоцена господствующее положение занял
человек, путь развития млекопитающих резко изменил свое направление. Палеонтологическая
летопись ранней фазы создает представление, что эволюция довольно часто
экспериментировала, временами неудачно, разрабатывала и опробовала различные
модели, выбирая те, которые могли бы выдержать конкуренцию и преуспеть в той
или иной природной среде. Во многих ранних моделях были обнаружены
недостатки, и они были одна за другой отвергнуты. Уцелевшие виды
просуществовали всю раннюю фазу и в ходе поздней фазы оформились и
развивались по вполне определенным направлениям. Многие из них существуют до
сих пор, занимая господствующее положение среди млекопитающих новейшего
времени.
Фото 48. Uintatherium,
травоядное млекопитающее массивного сложения, высотой около 1,5 м на фоне реконструированного
эоценового ландшафта. Его причудливо украшенный череп едва ли был полезен при
обороне или при нападении. Возможно, он привлекал особей противоположного
пола, а если так, то естественный отбор мог способствовать развитию этих
выростов Ранние
неспециализированные группы. Примитивный характер раннекайнозойских
млекопитающих (фото 48, рис. 55) объясняется двумя главными причинами.
Во-первых, эти млекопитающие происходили непосредственно от позднемеловых
видов, которые сами по себе были примитивными. Во-вторых, широко
распространенные в это время лесные ландшафты привели к сохранению архаичных
форм животных. Вероятно, большая часть раннекайнозойских
млекопитающих принадлежала к группе плацентарных. Как правило, они были
небольших или средних размеров (хотя один из них достигал величины
современного носорога). Ступня этих млекопитающих была плоской или почти
плоской (рис. 55, А), а у некоторых массивной, как у слона, и почти все они
имели пять пальцев. Среди них были как травоядные (большей частью питающиеся
побегами деревьев), так и хищники. Последние выделялись главным образом
формой зубов и примитивными когтями, не имея резких отличительных
особенностей общего облика. Некоторые из раннекайнозойских
млекопитающих, как, например, Uintatherium,
показанный на фото 48, обладали странными костными выростами на черепе, а
самцы этого вида имели также длинные верхние клыки.
Рис. 55. Четыре представителя ранних и примитивных
кайнозойских млекопитающих. A. Patriofelis, хищник.
Б. Phenacodus, небольшое травоядное. Один из его
родственников, возможно, был родоначальником линии лошадей. В. Coryphodon, травоядное массивного сложения, возможно,
обитавшее в болотах. Г. Barylambda, травоядное
длиной около 2,5 м Среда
обитания и пища. Один за другим примитивные виды млекопитающих вымирали или в
ходе эволюции становились более развитыми формами при определяющем влиянии
двух главных факторов - окружающей среды и пищи. В раннем кайнозое были
широко распространены леса, редколесья и саванны. В таких условиях листья
деревьев и кустарников составляли основную пищу растительноядных животных,
медленно передвигающихся от дерева к дереву. От врагов эти животные спасались
главным образом за счет "маскировки", поскольку большинство из них
не могло бегать на большие расстояния. Здесь встречались и хищники, и они
также прибегали к маскировке, выслеживая добычу. Длительного преследования
травоядных животных хищниками обычно не происходило. Напротив,
в степях, которые начали появляться в эоценовом периоде и все шире
распространялись в остальную часть кайнозоя, травоядные могли питаться только
травой и злаками. Такие млекопитающие должны были наклоняться за пищей к
самой поверхности земли. Им приходилось постоянно двигаться в поисках свежей
травы и воды и, таким образом, перемещаться на далекие расстояния. Им также
приходилось спасаться бегством от неизбежных хищников, так как спрятаться
здесь было негде. Их шансы на выживание могли увеличиться лишь за счет
усиленного размножения или объединения в большие группы или стада. Перед
хищниками вставала необходимость "развития" большой скорости, чтобы
преследовать добычу, так как они также не могли маскироваться, выслеживая
добычу. Мы
знаем, что некоторые млекопитающие всеядны. Например, современные медведи
едят плоды, ягоды, муравьев и рыбу. Они могут существовать в различных
условиях, обычно там, где есть реки и по крайней мере какие-нибудь деревья,
поскольку у них мало врагов и скорость передвижения не имеет для них большого
значения. Мы
вправе ожидать, что такие различия в условиях жизни млекопитающих и пищевом
рационе существенно повлияли на их форму тела и строение. Это становится
ясно, когда мы изучаем ряд ископаемых скелетов, полученных из достаточно
полного геологического разреза. Эволюция
конечностей и зубов. Ископаемые скелеты, собранные в различных отложениях
мезозоя, обнаруживают прогрессирующие изменения в строении, очевидно
связанные со средой обитания и с тем способом питания, который был обусловлен
этой средой. Млекопитающие, которые паслись в лесах и редколесьях, сохраняли
примитивную, довольно плоскую стопу, которая, как мы предполагаем, была
типична для мезозойских млекопитающих. У них сохранялись также неспециализированное
коренные зубы, служившие для пережевывания лиственной пищи. Постепенно ноги
их удлинялись, что отчасти позволяло им лучше спасаться от хищников, которые
постепенно становились более опасными. У
плотоядных животных, которые охотились и выслеживали травоядных, также
сначала сохранялась плоская стопа. Но понемногу их конечности приспособились
к быстрому бегу на короткие расстояния. Пятка (п) у
них отделилась от земли (рис. 56, Б), конечности стали длиннее и легче, и
мускулы бедра образовали пучок, необходимый, чтобы обеспечить быстрый рывок в
начале преследования. Хорошо развитые когти позволяли схватывать и валить
добычу. Заостренные передние зубы давали возможность кусать, а сильные мышцы
челюстей грызть добычу.
Рис. 56. Кости конечностей четырех различных видов
млекопитающих Конечности
и зубы травоядных млекопитающих также претерпевали изменения, хотя совсем
иначе. Так как пасущиеся животные должны были бегать, у них значительно
увеличилась длина и бедра и стопы; мускулы стали более сильными. Пятка
приподнялась так высоко, что стопа стала почти вертикально (рис. 56, В), а
когти на пальцах стали длиннее и толще. Параллельно с этим произошло удлинение
отдельных пальцев, так что другие перестали прикасаться к земле, постепенно
уменьшились и в конце концов совсем исчезли (рис. 57). Такое изменение
уменьшило вес стопы и способствовало увеличению скорости бега у травоядных
млекопитающих и дальности их перемещения. Эта эволюция, происходящая более
или менее одновременно у различных видов млекопитающих, может служить
свидетельством определяющего влияния среды. Однако вследствие дифференциации
среды изменение стопы зашло у отдельных видов млекопитающих дальше, чем у
других. У свиней "действуют" четыре пальца, у носорогов - три, у
верблюдов, оленей и крупного рогатого скота - два, у лошадей - один.
Рис. 57. Кости передних конечностей четырех родов
ископаемых лошадей. Через 50 миллионов лет эволюционного развития число
пальцев уменьшилось от четырех до одного, а общая длина конечности
увеличилась почти в 12 раз Этот
процесс занял долгое время. Например, лошади, впервые появившиеся в эоцене,
дошли до своего современного состояния (то есть стали иметь один палец) к
раннему плиоцену, а это значит в течение почти 60 миллионов лет. Если
поколения лошадей сменяются в среднем через четыре года, то этот процесс
охватил 15 миллионов поколений - 15 миллионов возможностей для изменений,
причем каждое поколение подвергалось воздействию среды, имевшему то же самое
направление развития. Если бы мы взяли 15 миллионов лошадей, по одной от
каждого поколения, и выстроили их друг за другом, начиная с небольшой
эоценовой лошади и кончая крупной современной, и они проходили бы мимо нас со
скоростью около 6,5 километра в час, то этот парад продолжался бы более
четырех месяцев. Наблюдая эту процессию в течение часа (в любой момент из
этих четырех месяцев), мы не увидели бы заметных изменений в облике животных.
Но к концу процессии вместо небольших, питающихся ветками животных высотой
около 30-40 сантиметров, передвигающихся, опираясь на четыре пальца, мы
увидели бы лошадей, опирающихся на один палец и имеющих высоту в холке до
полутора метров (рис. 58).
Рис. 58. Четыре вида лошадей, живших в различные
эпохи. Eohippus и Mesohippus
в действительности были еще меньше по отношению к более поздним лошадям, чем
это показано на рисунке В
ископаемых скелетах степных травоядных заметны и другие эволюционные изменения.
У них удлиняются челюсти, причем происходит разделение зубов на резцы,
расположенные спереди, и зубы, расположенные сзади и предназначенные для
жевания. Но так как степные травы содержат большое количество кремния, а
кроме того растительность степи часто покрыта пылью, серьезную проблему
представляло истирание коренных зубов мелкими твердыми частицами кремнезема.
Ход эволюции привел к видоизменению коренных зубов. У лошадей эти зубы
приобрели высокую коронку и складчатые образования эмали (рис. 59),
напоминающие модель Аппалачских гор в миниатюре. Эти складки включают твердый
слой эмали и прилегающий к ней более мягкий материал. При жевании пыльной
травы зубы снашиваются, но эмаль стирается в последнюю очередь и образует
выступы, придавая поверхности зуба шероховатость и облегчая перетирание
зеленой пищи.
Рис. 59. Коренные зубы млекопитающих. А. Низкий
коренной зуб нетравоядного млекопитающего. Б.
Коренной зуб травоядного млекопитающего, более высокий и имеющий складчатую
структуру; более мягкий материал быстрее истирается, а твердая эмаль (на
рисунке показана частой штриховкой) образует небольшие выступы В
отличие от описанных случаев далеко зашедшей специализации, некоторые
млекопитающие были очень слабо специализированы. Они не были ни травоядными,
ни плотоядными, могли есть и действительно ели все, что попадалось. Они не
оказывали четкого предпочтения ни лесу, ни степи, но жили в любом районе, где
была хорошая пища в достаточном количестве, будь то насекомые, ягоды или
плоды, рыба или мелкие млекопитающие. Они были искателями и скитальцами.
Типичными современными примерами таких животных являются медведи, еноты,
некоторые виды обезьян. Млекопитающим, ведущим такой образ жизни, не
требовалось вследствие условий жизни или пищевого рациона изменять форму
своих зубов и конечностей. Они навсегда сохранили примитивную плоскую стопу
(рис. 56, Г) и низкие коренные зубы (рис. 59 А). Эволюция
млекопитающих на суше, в море и в воздухе Если
проследить эволюционные линии всех ископаемых млекопитающих, как мы это уже
сделали с лошадьми, эти линии, разветвляясь, образуют дерево, подобное схеме
развития динозавров. Схема млекопитающих значительно сложнее, но на рисунке
60 дан ее упрощенный вариант, включающий только основные ветви. Однако даже
на этой упрощенной схеме видно, что большинство ветвей отделилось от главного
ствола в эоцене. История эволюционных изменений основных линий слишком
сложна, чтобы излагать ее полностью. Интересно, однако, рассмотреть хотя бы
немногие из исчезнувших ныне видов, принадлежавших к заканчивающимся тупиком
эволюционным линиям, не показанным на нашей схеме. Некоторые из них
изображены на рисунке 61. С нашей точки зрения, они выглядят странно. Но было
бы, однако, несправедливо считать эти виды неудачными только на том
основании, что они вымерли и, поэтому их облик непривычен для нас.
Рис. 60. Эволюционные линии млекопитающих В
отличие от странных животных, показанных на этом рисунке, крупные хищники
сохраняли относительную стабильность, не подвергаясь таким сильным
эволюционным изменениям, как лошади. В начале кайнозоя, как было установлено,
кошки подразделились на две группы - кошки, к которым относятся львы, тигры и
современные домашние кошки, и саблезубые кошки более массивного сложения. У
последних (фото 49) верхние клыки были длинными и уплощенными, а челюсти видоизменены
таким образом, что могли широко раскрываться. Прыгая на добычу, такая кошка
глубоко вонзала свои саблеобразные клыки, а затем вытаскивала их, оставляя
глубокие кровоточащие раны.
Рис. 61. Некоторые млекопитающие, обладавшие
приспособлениями, оказавшимися неудачными. A. Platybelodon,
млекопитающее, похожее на слона, у которого рот напоминал огромный совок; им
животное сгребало болотные растения. Миоцен. Б. Dinotherium
размером со слона, имел загнутую книзу нижнюю челюсть с клыками. Миоцен.
Рис. 61. Некоторые млекопитающие, обладавшие приспособлениями,
оказавшимися неудачными. В. Moropus, похожий на
лошадь, но более крупный, - травоядное животное с короткими задними ногами.
Его трехпалая стопа была снабжена когтями, возможно, служившими для
выкапывания растений. Миоцен. Г. Brontotherium,
травоядное длиной более 3,5 м. Олигоцен. Д. Syndyoceras,
родственный оленю, но с дополнительным рогом. Миоцен. Е. Epigaulus,
рогатый грызун с необычайно длинными когтями. Плиоцен К
олигоцену обе группы кошек так хорошо приспособились к своему хищному образу
жизни, что с тех пор они мало менялись. Саблезубый тигр вымер немногим более
10 000 лет назад, но мы не можем считать его неудачной моделью только на том
основании, что он вымер. Два или более видов его существовали на протяжении
более 30 миллионов лет. И мы не можем считать, что здесь эволюция зашла в
тупик. Действительно, вполне может быть, что саблезубые исчезли только
потому, что человек каменного века со своими усовершенствованными орудиями
охоты успешно боролся с ними за пищу. Как
мы уже видели, кайнозойские млекопитающие повторили ход развития рептилий -
подобно им, они увеличивали разнообразие видов и численность и достигли
господствующего положения. Как и мезозойские рептилии, они успешно проникли в
водную и воздушную среду. У мелких млекопитающих, живущих на деревьях,
развились крылья, состоящие из кожистой перепонки, и тем самым было положено
начало эволюционной линии летучих мышей. Крыло летучей мыши сконструировано
лучше, чем крыло летающей рептилии (рис. 52), но оно уступает крылу птицы по структуре,
а также потому, что на нем нет перьев. Может быть, это и есть главная причина
того, почему летучие мыши никогда не достигли такого успеха, как птицы.
Фото 49. Сцена времен олигоцена на территории
западной части современных Соединенных Штатов; саблезубая кошка (Hoplophoneus) убивает примитивную лошадь (Mesohippus). Реконструкция В
водной среде млекопитающие также повторили путь развивающейся группы
мезозойских рептилий. Не успела начаться эоценовая эпоха, как с суши в море
переселились наземные хищники, превратившиеся в примитивных китов с острыми
зубами. Возможно, они прошли промежуточную стадию, заходя на мелководье и
охотясь на рыбу. Один из этих видов развился позднее в настоящих китов,
которые разделились на несколько известных сейчас видов. Самое крупное из
ныне живущих морских млекопитающих, синий кит, по длине равен самому большому
из когда-либо существовавших динозавров, а по весу превосходит его более чем
вдвое. В
точности так же, как это произошло у их предшественников - рептилий за 200
миллионов лет до них, у всех морских млекопитающих тело удлинилось и
конечности превратились в плавники или ласты. Снова мы с поразительной
ясностью видим способность среды изменять форму и строение организмов.
Притягательная сила кайнозойских морей действовала не только на китов. В
эоцене одно из растительноядных млекопитающих, возможно состоявшее в
отдаленном родстве с предками слонов, также погрузилось в воду и
присоединилось к обитателям моря, основав линию, которая привела к
современным морским коровам и ламантинам. И в довершение к этому в миоцене хищник,
предположительно родственный кошачьим, присоединился к прежним обитателям
моря и дал линию современных тюленей. Дальние
миграции Палеонтологическая
летопись ясно свидетельствует о том, что млекопитающие неоднократно
мигрировали с одного континента на другой. Мы находим один и тот же род или
близкие роды на двух разобщенных массивах суши. Как могло произойти такое
расселение? Пыльца растений переносится по воздуху на тысячи километров, но
пылинки имеют микроскопические размеры. Насекомые заносятся ветром за сотни
миль водных пространств. Семена растений могут переноситься течениями через
Атлантический океан. Стволы деревьев также проплывают большие расстояния.
Известны случаи, когда змеи и крокодилы (и те и другие привычны к водной
среде) проплывали сотни километров между отдельными островами. Полярные
медведи проходят сотни километров по поверхности морских льдов. Но все это
частные случаи, которые не могут объяснить данные палеонтологической
летописи, свидетельствующие о том, что десятки видов млекопитающих размерами
от мыши до слона переходили с одного континента на другой. Такие
миграции нельзя объяснить с помощью гипотезы движения плит коры, при котором
происходит разъединение континентов и отторжение их частей далеко друг от
друга, потому что Атлантический океан существовал задолго до начала кайнозоя.
Что же касается возможности переплыть морское пространство, то очень мелкие
млекопитающие, возможно, и могли переплывать его на стволах деревьев, но для
большинства остальных животных единственный путь с одного континента на
другой (если не пользоваться целой флотилией ковчегов гораздо большего
размера, чем знаменитый ковчег Ноя) - сухопутные мосты. Обратимся к карте
континентов (рис. 16), на которой показаны также шельфы. На этой карте,
во-первых, видны узкие перемычки суши, существующие сейчас между двумя
Америками и между Европой и Африкой, и, во-вторых, мелководные участки, где
при довольно незначительных изменениях уровня океана относительно суши два
или более массивов суши могут оказаться соединенными (или почти соединенными
цепочкой островов между ними). Примером может служить связь Евразии с
Северной Америкой через Сибирь и Аляску и Евразии с Австралией через острова
Малайского архипелага. Таким образом, при небольших подвижках суши или
изменениях уровня моря могут возникнуть или мосты суши, или мелководья,
соединяющие между собой все континенты, кроме Антарктиды (отметим, что речь
идет лишь о последних 63 миллионах лет). Мост
между Аляской и Сибирью. Перемычка суши между Аляской и Сибирью может быть
восстановлена, если суша поднимется или уровень моря снизится всего лишь на
45 метров, Есть основания полагать, что здесь в течение большей части
кайнозоя существовал широкий мост суши. По нему происходило перемещение
различных млекопитающих в обоих направлениях. Вот почему среди ископаемых
млекопитающих Америки и Евразии так много близких форм и почему некоторые из
них почти совершенно идентичны. Мы, конечно, не можем считать, что эти
млекопитающие обладали таким "интеллектом", что могли "запланировать"
и осуществить это длинное путешествие в одну сторону. Они просто перемещались
в направлении, где была хорошая, удовлетворявшая их пища и где было не
слишком много конкурентов, претендующих на нее. Паразитирующие насекомые
переселялись вместе с травоядными животными, а крупные кошки и другие
плотоядные играли роль убийц и грабителей на большой дороге. Две
Америки. Иной была история Панамского перешейка, соединяющего сейчас две
Америки; она также оказала сильное влияние на развитие млекопитающих в
кайнозое. В палеоцене и раннем эоцене перешеек был сушей. Это позволило
некоторому количеству очень примитивных, неспециализированных видов
просочиться на южный континент. Затем позднее, в эоцене, связь между
континентами была прервана в результате опускания перешейка под воду; он
оставался погруженным в течение почти 50 миллионов лет, до конца плиоцена. В
течение этого времени примитивные млекопитающие (фото 50), в том числе
сумчатые, перекочевавшие в Южную Америку до того, как прервалось сообщение
между континентами, медленно развивались, защищенные от вторжения более
развитых видов из других мест. Но когда мост был восстановлен, огромное
количество развитых, современных млекопитающих (включая крупную кошку,
называемую ягуаром) нахлынуло сюда с севера и стало конкурировать с более
примитивными коренными обитателями. Один за другим местные виды исчезали, и у
нас есть основания полагать, что своей печальной участью они были в
значительной степени обязаны также первобытному человеку.
Фото 50. Необычайные южноамериканские млекопитающие,
обитавшие на территории Аргентины в плейстоцене. Слева - наземные ленивцы,
справа - глиптодонты (гигантские броненосцы) с панцирем, как у черепах.
Реконструкция Как
и на мосту, соединявшем Аляску и Сибирь, движение с одного американского
континента на другой происходило по двум направлениям, но поток, направленный
на север, был не столь уж велик. В Северную Америку проникли броненосцы,
наземные ленивцы и глиптодонты (гигантские броненосцы), причем первые из них
до сих пор обитают в Техасе. Сумчатые
Австралии. Каждый знает кенгуру - этот национальный символ Австралии.
Фактически, как и все другие млекопитающие Австралии, кенгуру относятся к
сумчатым млекопитающим. Благодаря своему примитивному строению они не могут
конкурировать с плацентарными млекопитающими, и, если бы последние смогли
появиться в Австралии в большом количестве, они, вероятно, давным-давно
вытеснили бы сумчатых. В Северной Америке, где сумчатые и плацентарные одно
время конкурировали между собой, все процветавшие ранее сумчатые исчезли, за
исключением представителей одной ветви, которая привела к опоссуму,
единственному сохранившемуся здесь до сих пор сумчатому. В
Австралии сумчатые жили гораздо лучше, и причиной этого была слабая
конкуренция со стороны других наземных животных. Среди современных животных
Австралии, помимо сумчатых, встречается только два вида однопроходных,
несколько плацентарных грызунов, похожих на крыс, летучие мыши, мелкие
рептилии и амфибии. Почти полное отсутствие плацентарных млекопитающих среди
местной фауны лучше всего объясняется изложенной ниже гипотезой, которая хотя
и не доказана, но очень правдоподобна. На карте (рис. 16) можно видеть, что
между Австралией и соседней с ней Азией существует обширное мелководье, на
котором так много островов, что проливы между ними нешироки. Предполагается,
что в самом конце мелового периода или в начале палеоцена примитивные
сумчатые попали в Австралию, "прыгая" с острова на остров, то есть
отдельные особи так или иначе случайно преодолевали узкие проливы между
островами. Они заселяли один остров, затем происходило новое случайное
переселение, и так продолжалось до тех пор, пока сумчатые не достигли Австралии.
В течение 20 или 30 миллионов лет ряд таких происшествий должен был произойти
неизбежно. Если
мы примем предположение, что австралийские сумчатые произошли от этих
путешественников, перебиравшихся с острова на остров, мы должны принять также
и гипотезу, что плацентарные млекопитающие могли осуществить это так же, как
и сумчатые, если не лучше. Возможно, что, когда Австралия в какой-то момент
до начала мелового периода отделилась от предположительно существовавшего
тогда древнего большого континента, показанного на рисунке 27, и стала
перемещаться к северу вместе с плитой земной коры, на ней уже жили ранние
формы сумчатых. Плацентарные млекопитающие в это время еще не появились. Но
какое бы объяснение мы ни приняли, результат остается одним: сумчатые в Австралии
были изолированы. Поэтому их развитие могло идти различными путями, избегая
губительной конкуренции со стороны более удачливых плацентарных
млекопитающих. Развитие сумчатых в основном заключалось в приспособлении к
различным типам природной среды, встречающимся в пределах обширного
австралийского континента. В пределах каждого типа среды сумчатые приобретали
такие формы, которые были лучше приспособлены к данному образу жизни, -
формы, напоминающие плацентарных млекопитающих, живущих в подобных условиях.
Так, например, в число современных сумчатых входят вомбат, бандикут, сумчатая
летяга, сумчатый (тасманийский) волк и сумчатая мышь. Эти животные могут быть
сопоставлены соответственно с плацентарными североамериканским сурком,
кроликом, белкой-летягой, волком и мышью. Эти сопоставления еще раз
показывают сильное влияние условий окружающей среды на ход эволюции; форма
тела животного и его способности вырабатываются в соответствии с образом
жизни, обусловленным внешними условиями, вне зависимости от того, каким был
исходный материал эволюции. Позднее,
уже в кайнозое, появились плацентарные животные, напоминающие крыс, а за ними
летучие мыши и змеи. Летучие мыши легко могли быть занесены с ветром во время
штормов; остальные могли приплыть на стволах деревьев. Они не представляли
сколько-нибудь серьезной угрозы с точки зрения борьбы за существование и, как
кажется, не повлияли сколько-нибудь существенно на развитие сумчатых. Жизни
сумчатых ничто не угрожало до тех пор, пока сравнительно недавно не появился
человек - охотник, и с ним другой охотник - собака. Прибытие европейцев в
Австралию имело катастрофические последствия. Сейчас сумчатые один за другим
исчезают. На
примере одного из животных, которых завез человек, - кролика (плацентарного)
- можно видеть, как происходит расселение новых видов животных на свободной
территории. В 1859 г. из Англии в Австралию были завезены две дюжины
кроликов. В течение следующих 69 лет потомство этих кроликов распространилось
почти по всей Австралии. На площади, составляющей приблизительно 1/3 всей
страны, в 1928 г. был сделан подсчет, показавший, что кроликов здесь
приблизительно полмиллиарда. Эта цифра помогает понять, как шло расселение
рептилий в мезозое. Заключение.
Таким образом, расселение наземных животных во всем мире очень близко к тому,
какое должно было бы быть, если бы существовали перемычки суши или цепочки
островов между северными континентами и не существовало никаких мостов, по
крайней мере в кайнозое, к югу от Австралии или Южной Америки. Эти два
континента представляли собой тупики, потому что на них обитают в основном
животные, принадлежащие к тупиковым линиям эволюции. Временами, на протяжении
50 миллионов лет или более, на них попадали млекопитающие, но дальше они
двигаться не могли. Мостов суши, соединявших их с Антарктидой, не
существовало. Благодаря отдаленности и низким температурам антарктического
континента животные попадали сюда только изредка и случайно. Даже сейчас
высшие животные представлены там только тюленями и пингвинами; и те и другие
могли попасть сюда и без всяких мостов. Значение
истории млекопитающих Кайнозойская
история млекопитающих далеко не такая длинная, как история мезозойских
рептилий, но для нас она гораздо важнее. Она прослежена на основе гораздо
большего количества ископаемых и представляет более разнообразные виды
животных. География и природные условия кайнозоя гораздо ближе к современным,
чем мезозойские. Таким образом, нам более знакома обстановка, в которой
развивались события. Наконец, млекопитающие принадлежат к той же группе, что
и мы сами, и потому представляют для нас большой интерес. В развитии
млекопитающих мы более ясно видим возникновение особенностей строения и
функций, которые существуют и в нашем теле. В
этой главе мы не останавливались на структурных и функциональных особенностях
организмов; мы даже не упомянули приматов, особую группу, к которой
принадлежим и мы сами, ограничившись указанием ее места в общей системе на
рисунке 60. Приматы для нас настолько важны, что они заслуживают особого
рассмотрения. После того как мы рассмотрим природные условия позднего
кайнозоя и климатическую обстат новку,
создавшую эти условия, мы обратимся в главе восемнадцатой к приматам и
посмотрим, к чему они пришли в результате своего развития. |